第三篇 医学蠕虫
第一节 概论
第二节 华支睾吸虫
第三节 布氏姜片虫
第四节 肝片形吸虫
第五节 卫氏并殖吸虫
第六节 斯氏狸殖吸虫
第七节 血吸虫
第八节 其它人体寄生吸虫
一、异形吸虫
二、棘口吸虫
蠕虫(helminth)为多细胞无脊椎动物,藉身体的肌肉收缩而作蠕形运动,故通称为蠕虫。在动物分类学史上,蠕虫曾被认为是独立的,具有特殊性的一类动物。但在分类学研究不断发展之后,人们发现蠕虫,实际上与人体有关系的包括扁形动物门(Phylum Platyhelminthes)、线形动物门(Phylum Nemathelminthes)和棘头动物门(Phylum Acanthocephala)所属的各种动物。因此在分类学上,蠕虫这个名称已无意义;但习惯上仍沿用此词。由蠕虫引起的疾病称为蠕虫病,具有重要意义的蠕虫种类几乎全部属于前两门。
第十四章 吸虫
第一节 概论
吸虫(trematode)属于扁形动物门的吸虫纲(Class Trematoda)。寄生人体的吸虫属于复殖目(Order Digenea),称为复殖吸虫(digenetic trematode)。复殖吸虫虽然种类繁多,形态各异,生活史复杂,但基本的结构(图14-1)和发育过程略同。

图14-1 复殖吸虫成虫形态构造示意图
形态与功能
大多数的复殖吸虫的成虫外观呈叶状或长舌状,两侧对称,背腹扁平,通常具口吸盘(oral sucker)与腹吸盘(acetabulum),内部结构如下:
体壁组织吸虫成虫体表有凹窝、凸起、皱褶、体棘、感觉乳突等,其形态、数量、分布等随虫种与虫体部位而异。体壁由体被(tegument)与肌肉层组成。中间为实质组织(parenchymal tissue)和埋在实质组织中的消化、生殖、排泄、神经系统等,缺体腔。体被为具有代谢活力的合胞体(syncytium),从外到内由外质膜(external plasma membrane)、基质(matrix)与基质膜(basal plasmamembrane)组成。基质膜之下为基层(basement layer)。肌肉层由外环肌(circular muscle)与内纵肌(longitudinal muscle)组成。体棘位于基质膜之上。感觉器位于基质中,有纤毛伸出体表,另一端有神经突(nerve process)与神经系统相通。此外还有内质网(endoplasmic reticulum)、核糖体(ribosome)、吞噬体(phagosome)、线粒体(mitochondrion)、高尔基复合体(golgi complex)和各种分泌小体(secretory body)等(图14-2)。体被具有保护虫体、吸收营养物质、感觉等生理功能。在结构上,体壁可随虫种、环境条件、发育阶段的不同而有差别,也可出现更新。结构的差别与更新通常也有生理功能上的意义。

图14-2 复殖吸虫成虫体壁结构示意图
消化系统 消化系统由口、前咽(prepharynx)、咽(pharynx)、食管(esophagus)和肠管(intestine)组成。肠管通常分为左右两个肠支(cecum)。肠壁
为单细胞层,其外周为肌细胞层所包围,肠细胞含有丰富的粗面内质网与线粒体,还常有高尔基复合体与各种包涵物,其胞质伸出具浆膜的长绒毛样褶,游离于肠腔,以扩大吸收面积。吸虫的消化系统具有消化和吸收功能。消化一般开始于肠管前部而在肠支中完成,主要是细胞外酶的作用。
生殖系统 人体吸虫除血吸虫外都具有雌雄两性的生殖器官,称为雌雄同体(hermaphrodite)。雄性生殖系统由睾丸(testis)、输出管(vas efferens)、输精管(vas deferens)、储精囊(seminal vesicle)、前列腺(prostatic gland)、射精管(ejaculatory duct)或阴茎(cirrus)、阴茎袋(cirrus pouch)等组成。雌性生殖由卵巢(ovary)、输卵管(oviduct)、卵模(ootype)、梅氏腺(Mehlis' gland)、受精囊(seminal receptacle)、劳氏管(Laurer's canal)、卵黄腺(vitellaria)、卵黄管(vitelline duct)、总卵黄管(common vitelline duct)、卵黄囊(vitellaria reservior)、子宫(uterus)、子宫末段(metraterm)等组成(图14-3、4)。雌、雄生殖系统的远端在生殖窦(genital atrium)会合,开口

图14-3 复殖吸虫成虫卵巢-卵巢模区结构

图14-4 复殖吸虫成虫生殖末段结构
于生殖孔(genital pore)。精子产生后从雄性生殖系统转入雌性生殖系统到达受精囊,卵的受精通常在输卵管。受精卵和从卵黄腺来的卵黄细胞排出的卵壳前体物质,在卵模内形成卵壳,以后进入子宫,经生殖孔排出。在复殖吸虫成虫结构系统中,生殖系统最发达,代谢最旺盛。由于成虫每日产卵数量很大,合成代谢与能量代谢都必须满足需要;因此,从外界摄取的营养物质的大部分最终都在本系统消耗。当然,进入虫体的有害物质,包括杀虫药,如无特殊的组织选择性,也同样会大部分积聚在本系统,常造成其正常生理功能首先被损害,表现为产卵数量的下降,甚至完全丧失生殖能力,最后虫体死亡。
排泄系统 吸虫的排泄系统由焰细胞(flame cell)、毛细管(capillary tubule)、集合管(collecting tubule)与排泄囊(excretory bladder)组成,经排泄孔(excretory pore)通体外(图14-5)。焰细胞与毛细管构成原肾(protonephron)单位。焰细胞的数目与排列各种吸虫可有不同,通常用焰细胞式(flame cell pattern)表示。焰细胞(图14-6)有细胞核、线粒体、内质网等;胞浆内有一束纤毛,每一纤毛有两根中央纤丝(fibril)与9根外周纤丝组成。纤毛颤动时很像火焰跳动,因而得名。纤毛的有节律的摆动可带动和保持排泄系统内部液体的流动,并可能形成相当高的过滤压,促使含有氨、尿素、尿酸等废物的排泄液排出虫体外。

图14-5 复殖吸虫成虫排泄系统

图14-6 焰细胞结构
神经系统 咽的两侧各有一神经节(ganglion),有背索(esophageal commissure)相联。神经节向前后各发出三条神经干(nerve trunk),分布于虫体的背面、腹面及侧面。向后神经干之间在不同水平上有横索(transverse commissure)。由神经干发出的神经感觉末梢到达口吸盘、咽、腹吸盘等器官,以及体壁外层的许多感觉器。神经系统有乙酰胆碱酯酶与丁酰胆碱酯酶的活动,神经节中有神经分泌细胞的存在,说明神经系统功能很活跃。
生活史
复殖吸虫的生活史都要经历有性世代(sexual generation)与无性世代(asexual generation)的交替。无性世代一般寄生在软体动物(中间宿主),通常是腹足类(gastropod),如螺蛳等。也可是斧足类(pelecypod),如蚌类。有性世代大多寄生在脊椎动物(终宿主)。复殖吸虫的生活史虽复杂,各种吸虫也有差别,但生活史的基本型则包括卵(ovum)、毛蚴(miracidium)、胞蚴(sporocyst)、雷蚴(redia)、尾蚴(cercaria)、囊蚴(encysted metacercaria)、后尾坳(囊内脱去尾部的虫体称后尾蚴metacercaria)与成虫(adult)。毛蚴侵入螺蛳的淋巴系统内发育变成为胞蚴。胞蚴通过体表摄取营养物质,其体内的胚细胞团经过分裂、发育形成多个雷蚴。雷蚴体内的胚细胞团再分化发育为多个子雷蚴或大量的尾蚴。有的吸虫,在寒冷季节,雷蚴不产生尾蚴而只产生雷蚴,可连续数代。此种变换生殖现象,是多胚繁殖(也称蚴体增殖)的吸虫长期适应环境的结果。绝大多数的尾蚴都有体部和尾部,但形态结构各异。尾蚴从螺体逸出后或侵入第二中间宿主体内或在物体的表面形成囊蚴。有些吸虫缺雷蚴期或有两代以上的雷蚴期,也可缺囊蚴期,而尾蚴直接侵入终宿主发育为成虫。侵入终宿主后童虫通常都经过移行,然后到达定居部位。一般认为移行中的虫体可能识别宿主的不断改变的连续的刺激,使绝大多数的虫体能够按一定的移行途径到达定居部位,而不适宜宿主则不能提供必要的生理信号,因而出现异常的个体发生移行(ontogenetic migration),导致发育迟缓、死亡或异常和异位寄生。复殖吸虫童虫移行时经历的和成虫寄生的器官、系统,其生化环境各异,可以设想必须具备有效的机制和生化代谢方式才能维持生命。各种吸虫从不同的寄生环境摄取的营养物质可能不同,但多以糖原而不以脂类作为能量贮存的一种形式,也缺乏脂类氧化所必需的酶。
生理
人体寄生的吸虫属复殖目,其生活史过程较复杂,既未丧失自生生活的某些特性,又能较广泛适应动物机体内的各种理化条件。这种广泛的适应性和迅速应变能力,是吸虫的重要生理特征之一。
吸虫的营养代谢,一般认为是以糖作为一种重要的能源,蛋白质与脂肪酸较不重要。吸虫吸收已糖主要是通过皮层,以被动扩散或以易化扩散方式进行,后者摄入速度比前者快;吸收后多以糖原形式贮存于虫体实质中。实验证明酵解酶类普遍存在于吸虫,加以许多吸虫的寄生期是处在氧压低的环境中,因此认为无氧糖酵解对吸虫尤为重要;但是糖酵解的整个过程有待进一步研究。氨基酸不是成虫能量主要来源,但胞蚴能分解利用较多的氨基酸。不同吸虫的氨基酸库差别较大,转氨酶种类也不相同,能利用的氨基酸亦各异。吸虫缺少脂类代谢。因而脂类积存于组织和排泄系统中。
吸虫寄居部位的含氧量差别较大,各种吸虫利用氧的途径、需氧的程度各异。氧溶解于吸虫皮层或肠道内壁而进入体内,被游离的血红蛋白携带和储存在各种组织中,复殖目吸虫生活史过程经历几个氧压很不相同的生活环境。例如虫卵、毛蚴和逸离螺体的尾蚴处在氧压高的环境;胞蚴、雷蚴和未成熟的尾蚴处在氧压低的环境;在物体表面结囊的后尾蚴和生活在血液中的成虫处在氧压高的环境,而宿主组织内的囊蚴和消化道中的成虫处在氧压低的环境。因而,在氧压变化的不同环境中生存,吸虫呼吸代谢也发生相应的变化。
分类
我国常见寄生人体的吸虫分类见表14-1。
表14-1 我国常见寄生人体吸虫的分类与其主要寄生部位
| 复殖目 Digenea |
后睾科 Opisthorchiidae |
支睾属 Clonorchis |
华支睾吸虫 C.sinensis |
肝胆管 |
| 异形科 Heterophyidae |
异形属 Heterophyes |
异形异形吸虫 H.heterophyes |
肠管 | |
| 片形科 Fasciolidae |
姜片属 Fasciolopsis |
布氏姜片吸虫 F.buski |
小肠 | |
| 片形属 Fasciola |
肝片吸虫 P.hepatica |
肝胆管 | ||
| 并殖科 Paragonimidae |
并殖属 Paragonimus |
卫氏并殖吸虫 P.westermani |
肺(或脑) | |
| 狸殖属 Pagumogonimus |
期氏狸殖吸虫 P.skrjabini |
皮下(或肝) | ||
| 裂体科 Schistosomatidae |
裂体属 Schistosoma |
日本裂体吸虫 S.japonioum |
门脉系统 | |
| 棘口科 Echinostomatidae |
棘隙属 Echinochasmus |
日本棘隙吸虫 E.japonicus |
小肠 |
第二节 华支睾吸虫
中华分支睾吸虫[Clonorchis sinensis(Cobbold,1875)Looss,1907]简称华支睾吸虫,又称肝吸虫。成虫寄生于肝的胆管内,可引起华支睾吸虫病(clonorchiasis),又称肝吸虫病。本虫于1874年首次在加尔各答一华侨的肝管内发现。曾于湖北江陵县先后在西汉古尸和战国楚墓古尸查见此种虫卵,证明华支睾吸虫病在我国流行至少已有2300年以上的历史。
形态
成虫(图14-7)体形狭长,背腹扁平,前端尖细,后端略钝,体表无棘。虫体大小一般为10~25mm×3~5mm。口吸盘略大于腹吸盘,后者位于虫体前端1/5处。消化道的前部有口、咽及短的食管,然后分叉为两肠支伸至虫体后端。睾丸前后排列于虫体后端1/3处,呈分支状,从睾丸各发出一支输出管,约有虫体的中部会合为输精管,向前逐渐膨大形成储精囊。储精囊接射精管开口于生殖腔。无阴茎和阴茎袋。卵巢边缘分叶,位于睾丸之前,受精囊在睾丸和卵巢之间,呈椭圆形。营氏管细长,弯曲,开口于虫体背面。卵黄腺滤泡状,分布于虫体两侧,从腹吸盘向下延至受精囊水平。输卵管的远端为卵模,周围为梅氏腺。子宫从卵模开始盘绕而上,开口于腹吸盘前缘的生殖腔。
卵在近子宫前端已渐成熟,排出的卵呈黄褐色,内有成熟的毛蚴。卵甚小,平均为29×17µm,形状似芝麻,一端较窄且有盖,盖周围的卵壳增厚、形成肩峰,另一端有小疣状突起(图14-7、14-8)。
生活史
成虫寄生于人或哺乳动物的胆管内。虫卵随胆汁进入消化道混于粪便排出,在水中被第一中间宿主淡水螺吞食后,在螺体消化道孵出毛蚴,穿过肠壁在螺体内发育,经历了胞蚴、雷蚴和尾蚴3个阶段。成熟的尾蚴从螺体逸出,遇到第二中间宿主淡水鱼类,则侵入鱼体内肌肉等组织发育为囊蚴。终宿主因食入含有囊蚴的鱼而被感染。囊蚴在十二指肠内脱囊。一般认为脱囊后的后尾蚴沿肝汁流动的逆方向移行,经胆总管至肝胆管,
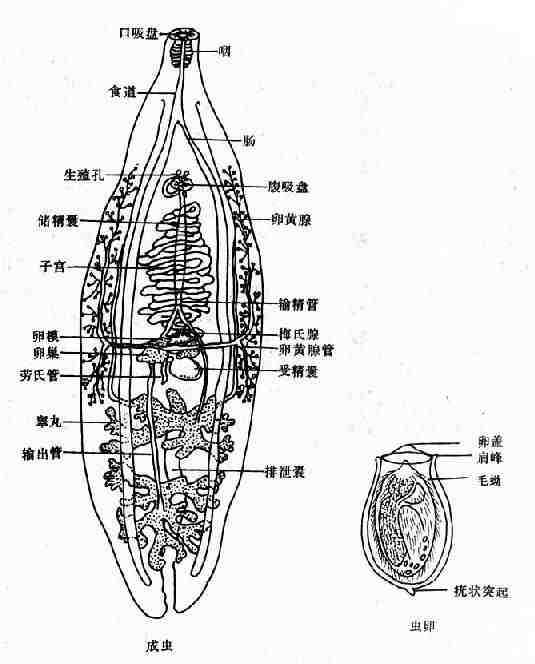
图14-7 华支睾吸虫成虫与虫卵
也可经血管或穿过肠壁经腹腔进入肝胆管内,通常在感染后1个月左右,发育为成虫(图14-9)。成虫在人体的寿命尚缺准确数据,一般认为有的可长达20~30年。
本虫对宿主的特异性要求并不严格。终宿主除肉食哺乳类动物外,在兔、豚鼠等食草动物也能正常发育。第一中间宿主淡水螺的种类很多,在我国至少有3科8种,常见的有纹沼螺(Parafossarulus striatulus),长角涵螺(Alocinma longicornis)和赤豆螺(傅氏豆螺(Bithynia fuchsianus)等。对第二中间宿主的选择更不强,国内已证实的淡水鱼宿主有12科39属68种。但从华支睾吸虫流行病学的角度看,养殖的淡水鲤科鱼类,如白鲩(草鱼)(Ctenopharyngodon idellus)、黑鲩(青鱼)(Mylopharyngodon aethiops)、鳊鱼(Hypophthalnichthys molitrix)、大头鱼(H.nobilis)、土鲮鱼(Labeo collaris及 L.kontius)鲤鱼(Cyprinus carpio)等特别重要。野生小型鱼类如麦穗鱼(Pseudorosbora parva)感染率较高,尚有克氏鲦鱼(Hemiculter kneri)等小鱼与传播儿童华支睾吸虫病有关。在台湾省日月潭地区,上述两种小鱼华支睾吸虫囊蚴的感染率甚至高达100%(Chen等.,1980)草鱼、鲫鱼的感染率也很高。除淡水鱼外,淡水虾如细足米虾(Caridna nilotica gracilipes)、巨掌沼虾(Macrobrachium superbum)等可有囊蚴寄生;甚至在某些特定条件下,尾蚴在螺体内也可发育为囊蚴。囊蚴寄生的部位,大部分在第二中间宿主的肌肉,但在鱼鳞和虾的甲壳表面也有发现(图14-9)。

图14-8 华支睾吸虫卵扫描电镜图(许世锷供图)
致病
本虫是厌氧性吸虫。在胆道中,成虫的营养和代谢还不很清楚。虫体组织化学研究及消化道内含物的分析结果显示消化道内有胆管上皮细胞、血液的各种成分和胆管分泌物等,但还不能认为这些物质是成虫的营养来源。虫体各组织中证实有碱性和酸性磷酸酶,可能与多糖、脂类、碱性蛋白质的分布有关。乙酰胆碱脂酶主要分布于神经系统内。
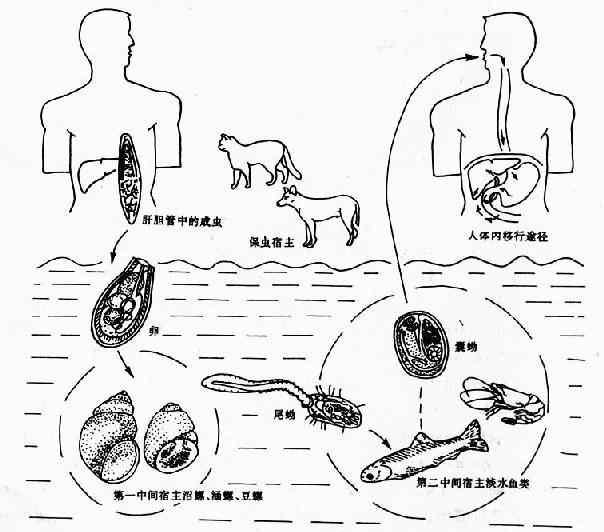
图14-9 华支睾吸虫生活史
华支睾吸虫病的危害性主要是患者的肝受损。虫体在胆道寄生时的代谢产物和机械刺激的结果。病变主要在肝的次级胆管。轻度感染或感染的初期病变并不明显。重度感染并经过相当长的时间后,胆管出现局限性的扩张,管壁增厚。大量的虫体可引起阻塞、胆汁滞留,如合并细菌感染可引起胆管炎和胆管肝炎。慢性感染可有大量的结缔纤维组织增生,附近的肝实质可用明显萎缩。目前一般认为虫卵、死亡的虫体及其碎片、脱落的胆道感染,破坏了胆道上皮的正常结构及功能,导致胆汗中细菌性β-葡萄糖醛酸苷酶活性升高,其结果有利于难溶性胆红素钙的形成;胆道分泌糖蛋白的增多,并附着于虫卵表面作为结石核心,起支架和粘附剂作用,促进胆红素钙的沉积,最后导致色素类结石(即肝内多发性结石)的出现(郭日波等,1990)。此外,国内外一些资料不断提示华支睾吸虫感染与胆管上皮癌、肝细胞癌的发生有一定关系。
临床症状以疲乏、上腹不适、消化不良、腹痛、腹泻、肝区隐痛、头晕等较为常见,但许多感染者并无明显症状。常见的体征有肝肿大,脾肿大较少见,偶见发育欠佳类似侏儒症者。严重感染者在晚期可造成肝硬变腹水,甚至死亡。
实验诊断
病原检查 检获虫卵是确诊的主要依据。但因虫卵小,粪便直接涂片法易于漏检,故多采用各种集卵法(如水洗离心沉淀法,乙醚沉淀法等)和十二指肠引流胆汗进行离心沉淀检查。但该虫卵与异形吸虫卵相似,不易鉴别。
免疫诊断 皮内试验、间接血凝试验、对流免疫电泳试验、酶联免疫吸附试验、间接荧光抗体试验等都曾试用于华支睾吸虫病的辅助诊断,但检测病人结果出入较大,且与其它消化道寄生虫感染(尤以吸虫类感染)有较明显的交叉反应,不能用作确诊,现仅作为流行病学调查初筛之用。随着免疫学新技术的发展和应用,如能找到理想的诊断用抗原,如纯化抗原、合成抗原等,, 百拇医药