脂联素及其信号传导
 |
 |
刘峰,美国华人糖尿病学会发起人之一,第一届学会主席。美国得克萨斯州立大学医学健康中心圣安东尼奥分校终身教授,美国国立卫生研究院(NIH)及美国糖尿病学会(ADA)项目评审委员会成员,中国国家自然科学基金海外评审专家,《美国生理学杂志》(内分泌学及代谢学)编委。目前主持的科研项目包括NIH-RO1基金资助项目2项、ADA资助项目1项及其他合作项目数项。独立从事基础研究12年,发表论文及综述50余篇。主要研究方向为胰岛素与脂联素的信号传导,肥胖、胰岛素抵抗、2型糖尿病及衰老的分子机制。
脂联素(adiponectin) 是一种新近发现由脂肪组织分泌、具有胰岛素增敏效应的激素。脂联素的含量和(或)多聚体的分布与糖尿病、动脉粥样硬化、炎症和肿瘤等的发生、发展及防治密切相关。脂联素除可通过自身信号通路实现其生理功能外,还可与其他信号通路相互作用,共同调节机体功能。
脂联素的分泌与结构
脂联素的功能及作用与其含量和多聚体的分布有重要关联
, 百拇医药
DsbA-L可能是调节脂联素合成、分泌和多聚体形成的重要功能蛋白之一
在机体中,脂联素的功能及作用与其含量和多聚体的分布有重要关系。虽然在肌组织、肝脏和心肌中发现有脂联素mRNA的表达,但一般认为,脂联素主要由白色脂肪组织中的脂肪细胞合成和分泌,也有报道表明,脂联素在棕色脂肪组织中能够表达且与产期葡萄糖及脂代谢有关。脂联素参与对多种生理和病理生理现象的调节,尤其是影响代谢性疾病、心血管疾病、肿瘤等的发生和发展。脂联素含量的降低见于肥胖、糖尿病、心血管疾病等,补充外源性脂联素或使用增加脂联素合成和分泌的药物可有效防止相关疾病的进展。脂联素含量的增加也见于某些严重疾病,如严重的心衰,其机制还不甚明了。
脂联素以低分子量、中分子量和高分子量3种多聚体形式存在于血浆或细胞中。研究表明,脂联素多聚体形式的分布,尤其是高分子量多聚体所占的比例决定其功能。笔者所在实验室最近通过酵母双杂交技术发现了一种与脂联素多聚体形成有关的重要调节蛋白,并将之命名为二硫键氧化还原酶A类似蛋白(DsbA-L)。在3T3-L1脂肪细胞中,DsbA-L过度表达可增加脂联素多聚体的形成。通过RNAi干扰技术抑制DsbA-L的表达,可导致高分子量脂联素显著减少,并伴随细胞水平和分泌的脂联素减少。在ob/ob鼠和肥胖患者的脂肪组织中,DsbA-L表达显著减低,同时脂联素含量和高分子量脂联素的比例也显著降低。这些结果表明,DsbA-L可能是调节脂联素合成、分泌和多聚体形成的重要功能蛋白之一。
, 百拇医药
脂联素信号传导通路
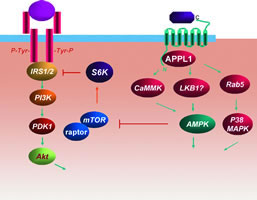
由APPL1介导的脂联素信号传导通路
可增加脂肪酸代谢及GLUT4膜转移
脂联素与其受体在膜外结合,激活下游的信号通路,从而增强细胞脂肪酸氧化和葡萄糖膜内转移。
脂联素受体 卡多瓦奇(Kadowaki)实验室最先报告了2个脂联素受体。AdipoR1存在于多种组织和细胞中,尤其在骨骼肌中表达较强。AdipoR2则主要在肝脏表达。
APPL1 我们运用AdipoR1的N-末端片段为诱饵,采用酵母双杂交技术对人胚胎大脑的cDNA 文库进行了分析,通过筛选得到一个与AdipoR1相结合的蛋白,并称之为APPL(adaptor protein containing pleckstrin homology domain, phosphotyrosine binding domain and leucine zipper motif)。 这也是首个被发现可介导脂联素信号的调节蛋白。在培养的肌细胞中,脂联素可增加APPL1与AdipoR1的结合,进而激活p38丝裂原活化蛋白激酶(p38 MAPK)和腺苷酸活化蛋白激酶(AMPK),并通过与Rab5(一种蛋白)结合介导脂联素刺激的葡萄糖转运子4(GLUT4)向细胞膜转移。
, http://www.100md.com
AMPK 在多数细胞中,脂联素可通过磷酸化AMPK位于活性区中的第172 位苏氨酸残基导致AMPK激活,进而增加线粒体的脂肪酸氧化。
MAPK 对于脂联素激活p38 MAPK的机制,目前了解不多。AMPK可能是p38 MAPK的上游激活激酶。我们的研究表明,无活性的Rab5过度表达能显著抑制脂联素诱导的p38 MAPK激活,提示脂联素激活p38 MAPK至少部分是通过APPL1与Rab5结合实现的。
■ 小结
脂联素的功能取决于其含量和多聚体的分布。DsbA-L在脂联素的合成、分泌和多聚体的形成中具有重要作用。脂联素通过与膜上的脂联素受体相结合,刺激APPL1与脂联素受体位于膜内的N-末端结合,从而介导AMPK和p38 MAPK激活。激活的AMPK和p38 MAPK可促进脂肪酸代谢及GLUT4的膜转移。APPL1和AMPK也是脂联素信号传导通路和胰岛素信号传导通路间的重要联系分子。通过这种相互作用,脂联素降低mTOR/S6K对IRS-1的负抑制,从而增加胰岛素的敏感性(图1)。
, 百拇医药
图1 脂联素信号传导及其与胰岛素信号传导的相互作用
胰岛素 胰岛素受体 脂联素受体 脂联素
葡萄糖摄取等 脂肪酸氧化、葡萄糖摄取等
通信联系(Cross-talk)
脂联素信号传导通路与胰岛素信号传导通路通过
APPL1及AMPK等信号分子相互联系
我们最近的研究表明,脂联素信号传导可通过直接通路影响胰岛素信号传导通路,从而增强胰岛素敏感性。胰岛素信号传导由两个重要的信号通路组成:磷脂酰肌醇3-激酶(PI3K)和MAPK通路。胰岛素通过与细胞膜上的胰岛素受体结合,使胰岛素受体自磷酸化以及包括胰岛素受体底物1(IRS-1)在内的某些底物磷酸化。IRS-1酪氨酸残基磷酸化后,顺序激活PI3K通路中的丝氨酸/苏氨酸蛋白激酶,如3-磷酸肌醇依赖性蛋白激酶-1(PDK-1)、蛋白激酶B(PKB)等,其中PKB在胰岛素调节的葡萄糖代谢中具有重要意义,其活性降低可见于2型糖尿病等。
, 百拇医药
脂联素具有增强胰岛素敏感性的作用。给2型糖尿病和胰岛素抵抗(IR)动物补充脂联素可增强IRS-1和Akt的磷酸化,而缺乏脂联素的小鼠表现出严重IR,说明脂联素信号通路和胰岛素信号通路间存在相互联系(cross-talk)。此种联系可能主要涉及以下几个信号分子:
APPL1 APPL1与脂联素受体结合,激活脂联素下游效应分子,如AMPK和p38 MAPK,在脂联素的胰岛素增敏效应和促脂肪酸氧化中起重要作用。APPL1也可能直接与胰岛素受体及下游信号分子结合,增强胰岛素信号传导。
AMPK 脂联素信号通路通过激活AMPK降低S6激酶(S6K)活性,从而减小对IRS-1的负调节作用,增强PI3K途径。在内皮细胞中脂联素促血管再生的作用可能是AMPK、PI3K、Akt及eNOS介导的。
其他分子 如NFB、C-Jun 氨基末端激酶(JNK)、mTOR 和S6K。激活的NFB可通过炎症因子介导IR。脂联素可能通过对NFκB的抑制而增加胰岛素敏感性。JNK可通过抑制IRS-1酪氨酸磷酸化及促进IRS-1丝氨酸磷酸化等多种途径导致IR和2型糖尿病。同时,JNK激活可导致脂联素分泌水平显著降低。与JNK活化相似,激活mTOR和S6K也促进IRS-1丝氨酸磷酸化,抑制其酪氨酸磷酸化,从而影响胰岛素信号传导。脂联素通过激活AMPK抑制mTOR/S6K活性,进而增强胰岛素信号传导。, http://www.100md.com