腺病毒亚属C型主要晚期基因三联先导序列对基因表达的调节
作者:黄文林 乔泰东 刘志国 樊代明
单位:美国普林斯顿大学分子生物学系(黄文林);第四军医大学全军消化病研究所(乔泰东、刘志国、樊代明)
关键词:腺病毒科感染;肌动蛋白类;信使核糖核酸
中华医学杂志/981004 【摘要】 目的 观察腺病毒主要晚期三联先导序列(tripartite leader sequence, tpl)在腺病毒C型感染细胞晚期对病毒mRNA转运的影响。方法 用构建的激素诱导型腺病毒重组体MA及MtplA(含有腺病毒主要晚期先导序列)感染细胞,采用Northern印迹、引物延伸及脉冲示踪等方法比较感染细胞中特异性报告基因β-肌动蛋白mRNA的转送特征。结果 尽管两种报告基因转录产物的合成速率相同,但在含有tpl的MtplA腺病毒感染的细胞中,胞浆内报道基因mRNA浓度更高。结论 对细胞核与细胞质之间MA和MtplA的新合成和稳态RNA的分析表明,tpl并不能增加病毒mRNA在胞质中的稳定性,但可促进病毒mRNA从胞核到胞质的转送。
, 百拇医药
Expression regulation of major late promoter tripartite leader sequence in adenovirus late infection
Huang Wenlin*, Qiao Taidong, Liu Zhiguo, et al. *
Department of moleculer Biology, Princeton University, USA.
【Abstract】 Objective To investigate the influence of adenovirus major late tripartite leader sequence on specific exportation of virus mRNA in the late phase of adenovirus infection.Methods We constructed a marked, human beta-actin minigene under the control of the glucocorticoid- inducible enhancer and promoter of mouse mammary tumor virus, and inserted it into the left end of the adenovirus type V genome. The promoter gene, designated MA, as well as of a sibling, which differed only in inclusion of a cDNA copy of the adenovirus major late tripartite leader sequence upstream of beta-actin sequences termed MtplA, in recombinant virus-infected cells was strictly dependent on addition of deoxymethesone to the medium. Nuclear and cytoplasmic reporter RNA species were compared by Northern blotting, primer extension and pulse-chase assay.Results The high concentrations of newly-synthesized RNA were accumulated in cytoplasm when tripartite leader sequence was present in reporter RNA, despite the equal rates of transcription of the two reporter genes. Nuclear and cytoplasmic reporter RNA species compared by Northern blotting, primer extension, and pulse-chase provided no evidence for altered processing induced by tripartite leader sequence. Conclusion The tripartite leader sequence, known to facilitate translation of mRNA during the late phase of adenovirus infection, can also modulate mRNA export from the nucleus.
, 百拇医药
【Key words】 Adenoviridae infections Actin mRNA
(Natl Med J China, 1998, 78:733-737)
我们利用人β-肌动蛋白(34个寡核苷酸)作为报告基因,以含有或缺失腺病毒Ⅴ型主要晚期启动子三联先导序列(tripartite leader sequence, tpl)的鼠乳腺瘤病毒启动子(MMTV)构建成MA或MtplA重组病毒[1,2]。这种激素诱导型腺病毒重组体在感染细胞后,可在感染细胞中观察到特异性报告mRNA的转送特征。为了更加精确地分析选择性转运mRNA对于感染晚期转录的影响,我们观察了腺病毒主要晚期tpl对病毒mRNA转运效率的影响
材料与方法
一、新合成RNA的分析
, 百拇医药
用30噬斑形成单位(pfu)/细胞的Ad5/Mtpl A和Ad5/MA感染293细胞,在感染后的10~14小时内加入3 mmol/L的地塞米松。1小时后,用PBS冲洗细胞2次,然后在37 ℃下孵育1~3小时,再用7.4 MBq(200 μCi)/ml 3 H-尿嘧啶[1468.9GBq(39.70 Ci)/mmol,NEN-Dupont公司)标记15分钟,在未标记尿嘧啶的溶液中追加10~15分钟,然后分离胞浆和胞核成分。胞浆部分在37 ℃用10 μg/ml蛋白激酶K孵育30分钟,随后用酚:氯仿抽提胞浆RNA 2次,乙醇沉淀。将细胞核部分溶解于含有20 mmol/L NaCl的4 mol/L异硫氰酸胍的40 mmol/L Tris-HCl (pH7.4)液中。离心12~15小时,可使RNA通过5.7 mol/L CsCl沉淀下来。将沉淀重悬于蒸馏水中,用含0.25 mol/L NaCl和0.005 mol/L EDTA的10 mmol/L Tris HCl (pH7.4)液透析。用3H标记的RNA与含有病毒和宿主细胞基因的质粒DNA杂交,同时也使之与SpeI寡核苷酸链杂交。由于Hsp 70及病毒主要晚期表达基因L2质粒可在293细胞中持续表达,主要使用一个minifold Ⅱ印迹转移装置,就可将变性的线性质粒DNA转移到膜上。在转移之前,需将SpeI寡核苷酸接点非编码链与鱼精DNA混合,经紫外光照射与膜交联后,在0.125 mol/L磷酸钠缓冲液中与3 H-尿嘧啶标记的胞浆和胞核RNA杂交。该缓冲液由50%甲酰胺、0.25 mol/L NaCl、体积分数为5%的SDS、体积分数为10%的PEG8000和1 mmol/L EDTA组成。杂交温度为55 ℃,时间为12~16小时。洗膜、测定杂交到不同目的DNA上的RNA的放射性[2]。
, 百拇医药
二、Northern杂交法
按前述方法感染细胞,分离细胞核和细胞浆RNA,通过寡脱氧核苷酸[Oligo(dT)]结合的磁珠可与含Poly(A)的RNA结合的特性,分离含有Poly(A)和缺少Poly(A)的RNA。胞核和胞浆RNA乙醛化后 在1.2%的琼脂糖凝胶上电泳。电泳缓冲液为10 mmol/L NaH2PO4(pH7.0)液。然后将RNA电转移到杂交膜上(美国Dupont公司),再将膜与SpeI寡核苷酸接点的非编码链,或一段与5′末端标记的与MMTV转录单位+28~+52碱基互补的寡核苷酸链杂交。利用T4多核苷酸激酶和[γ-32P]-ATP[111 GBq(3Ci)/mmol,NEN-Dupont公司]标记MMTV及β-肌动蛋白探针,然后洗膜,使用分子动态磷光计数仪检测杂交信号。
三、体外连续转录
从Ad5/MA和Ad5/MtplA感染的293细胞中分离细胞核成分,并将其悬于含有体积分数为40%的甘油、5 mmol/L MgCl2和0.1 mmol/L EDTA的50 mmol/L的Tris-HCl(pH8.3)中,计数浓度为108个核/ml。在30 ℃下延长起始RNA 30分钟,纯化标记RNA。标记的RNA与结合在尼龙膜上的Spel接头的寡核苷酸(pH 2.3)和PS73-ML质粒DNA杂交。杂交后冲洗、放射自显影方法同前。用闪烁计数方法测定膜相应部位杂交到不同DNA上的RNA数量。
, 百拇医药
四、β-肌动蛋白微基因转录本5′末端作图
由图1可见MA与MtplA基因核内和胞浆中5′端转录本。通过引物延伸法做图,该引物为与MMTV转录单位+28~+52位互补的片段。
五、加工后β-肌动蛋白微基因转录本的分析
分离纯化Ad5/MA或Ad5/MtplA感染细胞的胞浆和胞核RNA,以及分离含Poly(A)和不含Poly(A)成分的步骤如前所述。用逆转录聚合酶链反应(RT-PCR)法分离RNA,得到的扩增产物为杂交到人β-肌动蛋白微基因上游Poly(A)位或与Ad5基因4014-4055结合的引物延伸出来的片段。后者的序列位于重组病毒β-肌动蛋白小基因下游的E1B基因中(图1)。逆转录反应体系里含有200 U/μl的逆转录酶。PCR反应体系为:2.5 U的Taq聚合酶,2 mmol/L MgCl2,100 μmol/L dNTPs或100 μmol/L dATP、dGTP和dTTP,以及1 μmol/L dCTP、370 kBq(10 μCi)[α-
, 百拇医药
32 p]-dCTP(NEN-Dupont公司),与MMTV转录单位+21~+44位互补的3′引物,与3′端ML三联先导序列互补的5′引物和3′β-肌动蛋白引物。60个循环后,酚-氯仿抽提PCR产物,2倍体积乙醇沉淀,然后在1%琼脂糖凝胶上电泳,电泳缓冲液为TAE,用溴化乙淀染色或待凝胶干燥后,用放射自显影测定。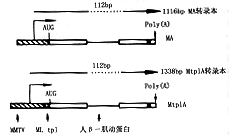
Ad5/MA和Ad5/Mtpl A的区别为后者在MMTV启动子与β-肌动蛋白基因间有一段主要晚期三联先导序列的导入,折线箭头为MMTV LTR启动子未剪割RNAs的启始,虚线表示人β-肌动蛋白基因的mRNA图1 激素诱导型β-肌动蛋白报导基因病毒
重组结构示意图
结果
一、Northern杂交检测报告基因产物
, 百拇医药
分离感染晚期细胞核及细胞浆RNA比较试验发现,带有tpl顺序的报告RNA更有效地聚积到胞浆,这可能是RNA加工、转送至胞浆的速率不同的结果。Northern杂交测定经地塞米松处理由MA和MtplA感染细胞的RNA,用插入到β-肌动蛋白微基因的寡核苷酸作分子探针,杂交结果显示一条与MA和mtplA报告基因剪接转录物的长度相吻合的条带,而且两种病毒报告基因RNA在胞浆中的分布不同。胞浆中的MA RNA是MtplA RNA的1/3,与此相反,MA RNA在胞核中的浓度明显大于MtplA RNA在胞核中分布。腺病毒报告基因特异性寡核苷酸及MMTV探针杂交显示的结果一致(图2,3)。
来自于Ads/MA感染Poly(A)细胞浆RNA及细胞核RNA(N)(3,5号样),Ad5/Mtp1A(2,4号样)感染细胞RNA,野生型Ad5(6号样)感染细胞RNA,与报告基因插入特异性的Spel寡核苷酸杂交图2 表达基因RNA杂交分析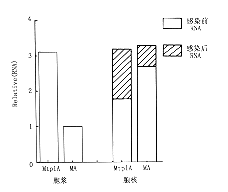
, 百拇医药
图3 分子动力磷构象分析(Molecular Dynamics Phosphorimager)
测定分离的杂交片段,得出两组报告基因感染细胞后
细胞浆及细胞核RNA分布
二、脉冲示踪标记法
为了更精确地评估MA和MtplA RNA在胞浆中的稳定性对胞质浓度差异的作用,采用脉冲示踪标记法检测新合成的报告NRA在核/浆中的比例。结果发现,感染晚期,[3H]-Uridin标记经地塞米松处理的293细胞,在标记15分钟,示踪20分钟,与其相关RNA杂交后进行报告RNA、Hsp70和L2病毒RNA浓度分布的比较,新合成的Hsp RNA和L2病毒RNA在核/浆分布与预料相同:病毒晚期mRNA高效地进入胞浆,而Hsp70 RNA的输出受抑制。细胞质中观察到新合成带有tpl序列的报告基因RNA浓度大于不含tpl序列报告的RNA浓度。而细胞核中这两种报告RNA的浓度刚好相反,它们的RNA总浓度相同。带tpl序列的RNA进入胞浆增加表明转运速率高。用短时间脉冲示踪可有效地排除因时间和实验温度、条件所引起的数据误差,并证明带有tpl序列的报告RNA进入胞浆的效率是缺失tpl序列RNA的2.4~4.1倍(附表,图4~8)。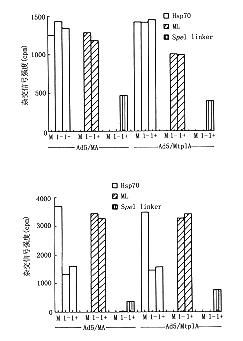
, 百拇医药
在同样的293细胞密度上用30 pfu/细胞的两种病毒(Ad/MA,Ad/Mtp1A)感染细胞,10小时后经地塞米松诱导,设感染对照(M),诱导(1+),无诱导(1-),14小时后,收集细胞核提取物作体外继续转录试验(图4).另一份感染细胞经[3H]-尿嘧啶进行示踪标记(图5)图4、5 β-肌动蛋白报告基因经诱导后在Ad/MA
和Ad/Mtp1A感染293细胞中的表达
三、腺病毒报告基因RNA的加工
引物延伸两种报告基因,在MMTV转录启始点+27~+52合成互补寡核苷酸片段,引物扩增的两种重组病毒都产生52个核苷酸的cDNA。Northern杂交和RT-PCR都没有发现带有病毒E1B序列的重组病毒特异性RNA,证实MA和MtplA的转录是独立系统,两种报告基因的转录启始不受残留的E1A增强子影响(图2,3,9)。
, http://www.100md.com
附表 重组腺病毒感染细胞后新合成RNA的有效转送率(%) 组别
新合成的细胞浆/细胞核RNA
HSP70
ML-L2
β-actin
Reporter
1.对照细胞
3.1
ND
ND
Ad5/MA+Dex,11h
0.3
, 百拇医药
8.5
0.1
Ad5/MtplA+Dex,11h
0.3
8.2
0.5
2.对照细胞
3.0
ND
ND
Ad5/MA+Dex,11h
0.5
2.5
, http://www.100md.com
0.3
Ad5/MtplA+Dex,11h
-
2.3
1.0
3.对照细胞
9.7
ND
ND
Ad5/MA+Dex,11h
0.4
11.7
0.1
, 百拇医药
Ad5/Mtp1A+Dex,11h
0.4
12.4
0.4
注:1.新合成Hsp70、ML-L2和β-肌动蛋白报告RNA在细胞质与细胞核中分布的定量,数字经图6~8分析获得,2.ND=检测不到,-=没检测;Dex=地塞米松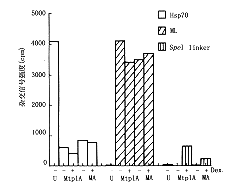
图6 两种重组腺病毒感染细胞后细胞浆RNA分布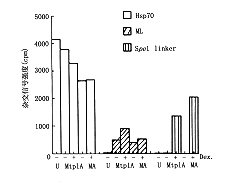
图7 两种重组腺病毒感染细胞后细胞核RNA分布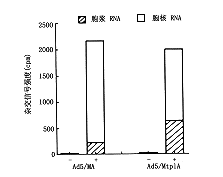
, 百拇医药
图8 两种重组腺病毒感染细胞后细胞核与细胞浆新合
成RNA分布
四、RT-PCR测定报告基因RNA长度
比较MA和MtplA RNA反转录产物的长度,发现MtplA的RNA比相应的MA RNA长150~170个碱基,恰好和预期的tpl序列长短(164)相吻合。并与剪接后去除内含子6的报告基因RNA的长度完全一致,这说明从报告转录物中剪接了内含子(图1,9)。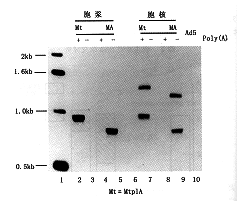
RNA经Ad5/MA和Ad5/mtplA感染,于地塞米松诱导后提取、纯化,应用人β-肌动蛋白4024~4055互补引物及5′+22/44引物.Ad5/MA RNA(4,8);Ad5/mtplA RNA(2,6)图9 RT-PCR测定腺病毒报告基因转录体
, http://www.100md.com
讨论
本研究利用腺病毒报告基因能被激素诱导等生物特性来研究腺病毒感染细胞后,其mRNA在转录后加工中,从胞核向胞浆的转运机制。报告基因的转录严格依赖于外源性激素,通过增强子上GRES激活MMTV转录调控区开始转录[1]。这证明报告基因的转录活性不受相邻腺病毒序列(如上游的E1A增强子)的影响。去除地塞米松后,报告基因可继续转录几小时。这是研究腺病毒感染细胞后转录的激活、转录物加工及核膜转运的理想模型。
在两株腺病毒感染细胞晚期,被地塞米松激活的报告基因转录物都是以剪接后产物形式进入细胞质,说明这些细胞浆中的MA和MtplA RNA是被特异性输出的。MA基因不带任何病毒序列,转录后外运到胞浆的量明显低于MtplA RNA,同时细胞本身的mRNA如β-肌动蛋白、热休克蛋白转送受抑制(图4,5,6,7,8,附表)。
主要晚期三联先导序列可能控制mRNA转录后多个代谢步骤,包括mRNA从胞核转送,胞浆中的稳定性调控及翻译过程[3]。tpl增强输出RNA的最直接机制可能是在感染细胞通过特异地识别某些病毒或细胞介导蛋白,以协助RNA进入核膜输出通道。tpl的作用可能类似于HIV-1 mRNA RRE。这个HIV-1 Rev可识别转录后剪接和未剪接的病毒RNA位点而协助向核外转运[4]。最可能与腺病毒tpl特异结合的蛋白是E1B-55 000和E4-34 000蛋白。然而,这些病毒早期蛋白允许选择输出无tpl序列的RNA和某些细胞mRNA,这提示它们不能特异地识别tpl“输出”信号,表明可能还有其它病毒或细胞蛋白参与tpl的输出功能。因为tpl是首次应用组装病毒方式发现其具有在细胞内转运RNA的能力的病毒序列,新发现的任何特异性识别核蛋白都会提供关于与RNA输出和调节分子机制有关的重要线索。
, 百拇医药
本文在文稿及图片的整理工作中得到吴静、尹芳、曹晓燕等的帮助,特此致谢
参考文献
1 Berkner KL, Sharp PA. Effect of tripartite leader on synthesis of a non-viral protein in an adenovirus 5 recombinant. Nuc Acids Res, 1985, 13:841-857.
2 Cullen BR, Malim MH. The HIV-1 Rev protein: prototype of a novel class of eukaryotic post-transcriptional regulators. TIBS, 1991, 16:346-350.
3 Derget SM, Moore C, Sharp PA. Spliced segments at the 5′ terminus of adenovirus 2 late mRNA. Proc Natl Acad Sci USA, 1997, 74:3171-3175.
4 Huang WL, Pruzan R, Flint SJ. In vitro transcription from the adenovirus E2 early promoter by RNA polymerase Ⅲ, Proc Natl Acad Sci USA, 1994, 91:1265-1269.
(收稿:1998-01-05 修回:1998-06-30), http://www.100md.com
单位:美国普林斯顿大学分子生物学系(黄文林);第四军医大学全军消化病研究所(乔泰东、刘志国、樊代明)
关键词:腺病毒科感染;肌动蛋白类;信使核糖核酸
中华医学杂志/981004 【摘要】 目的 观察腺病毒主要晚期三联先导序列(tripartite leader sequence, tpl)在腺病毒C型感染细胞晚期对病毒mRNA转运的影响。方法 用构建的激素诱导型腺病毒重组体MA及MtplA(含有腺病毒主要晚期先导序列)感染细胞,采用Northern印迹、引物延伸及脉冲示踪等方法比较感染细胞中特异性报告基因β-肌动蛋白mRNA的转送特征。结果 尽管两种报告基因转录产物的合成速率相同,但在含有tpl的MtplA腺病毒感染的细胞中,胞浆内报道基因mRNA浓度更高。结论 对细胞核与细胞质之间MA和MtplA的新合成和稳态RNA的分析表明,tpl并不能增加病毒mRNA在胞质中的稳定性,但可促进病毒mRNA从胞核到胞质的转送。
, 百拇医药
Expression regulation of major late promoter tripartite leader sequence in adenovirus late infection
Huang Wenlin*, Qiao Taidong, Liu Zhiguo, et al. *
Department of moleculer Biology, Princeton University, USA.
【Abstract】 Objective To investigate the influence of adenovirus major late tripartite leader sequence on specific exportation of virus mRNA in the late phase of adenovirus infection.Methods We constructed a marked, human beta-actin minigene under the control of the glucocorticoid- inducible enhancer and promoter of mouse mammary tumor virus, and inserted it into the left end of the adenovirus type V genome. The promoter gene, designated MA, as well as of a sibling, which differed only in inclusion of a cDNA copy of the adenovirus major late tripartite leader sequence upstream of beta-actin sequences termed MtplA, in recombinant virus-infected cells was strictly dependent on addition of deoxymethesone to the medium. Nuclear and cytoplasmic reporter RNA species were compared by Northern blotting, primer extension and pulse-chase assay.Results The high concentrations of newly-synthesized RNA were accumulated in cytoplasm when tripartite leader sequence was present in reporter RNA, despite the equal rates of transcription of the two reporter genes. Nuclear and cytoplasmic reporter RNA species compared by Northern blotting, primer extension, and pulse-chase provided no evidence for altered processing induced by tripartite leader sequence. Conclusion The tripartite leader sequence, known to facilitate translation of mRNA during the late phase of adenovirus infection, can also modulate mRNA export from the nucleus.
, 百拇医药
【Key words】 Adenoviridae infections Actin mRNA
(Natl Med J China, 1998, 78:733-737)
我们利用人β-肌动蛋白(34个寡核苷酸)作为报告基因,以含有或缺失腺病毒Ⅴ型主要晚期启动子三联先导序列(tripartite leader sequence, tpl)的鼠乳腺瘤病毒启动子(MMTV)构建成MA或MtplA重组病毒[1,2]。这种激素诱导型腺病毒重组体在感染细胞后,可在感染细胞中观察到特异性报告mRNA的转送特征。为了更加精确地分析选择性转运mRNA对于感染晚期转录的影响,我们观察了腺病毒主要晚期tpl对病毒mRNA转运效率的影响
材料与方法
一、新合成RNA的分析
, 百拇医药
用30噬斑形成单位(pfu)/细胞的Ad5/Mtpl A和Ad5/MA感染293细胞,在感染后的10~14小时内加入3 mmol/L的地塞米松。1小时后,用PBS冲洗细胞2次,然后在37 ℃下孵育1~3小时,再用7.4 MBq(200 μCi)/ml 3 H-尿嘧啶[1468.9GBq(39.70 Ci)/mmol,NEN-Dupont公司)标记15分钟,在未标记尿嘧啶的溶液中追加10~15分钟,然后分离胞浆和胞核成分。胞浆部分在37 ℃用10 μg/ml蛋白激酶K孵育30分钟,随后用酚:氯仿抽提胞浆RNA 2次,乙醇沉淀。将细胞核部分溶解于含有20 mmol/L NaCl的4 mol/L异硫氰酸胍的40 mmol/L Tris-HCl (pH7.4)液中。离心12~15小时,可使RNA通过5.7 mol/L CsCl沉淀下来。将沉淀重悬于蒸馏水中,用含0.25 mol/L NaCl和0.005 mol/L EDTA的10 mmol/L Tris HCl (pH7.4)液透析。用3H标记的RNA与含有病毒和宿主细胞基因的质粒DNA杂交,同时也使之与SpeI寡核苷酸链杂交。由于Hsp 70及病毒主要晚期表达基因L2质粒可在293细胞中持续表达,主要使用一个minifold Ⅱ印迹转移装置,就可将变性的线性质粒DNA转移到膜上。在转移之前,需将SpeI寡核苷酸接点非编码链与鱼精DNA混合,经紫外光照射与膜交联后,在0.125 mol/L磷酸钠缓冲液中与3 H-尿嘧啶标记的胞浆和胞核RNA杂交。该缓冲液由50%甲酰胺、0.25 mol/L NaCl、体积分数为5%的SDS、体积分数为10%的PEG8000和1 mmol/L EDTA组成。杂交温度为55 ℃,时间为12~16小时。洗膜、测定杂交到不同目的DNA上的RNA的放射性[2]。
, 百拇医药
二、Northern杂交法
按前述方法感染细胞,分离细胞核和细胞浆RNA,通过寡脱氧核苷酸[Oligo(dT)]结合的磁珠可与含Poly(A)的RNA结合的特性,分离含有Poly(A)和缺少Poly(A)的RNA。胞核和胞浆RNA乙醛化后 在1.2%的琼脂糖凝胶上电泳。电泳缓冲液为10 mmol/L NaH2PO4(pH7.0)液。然后将RNA电转移到杂交膜上(美国Dupont公司),再将膜与SpeI寡核苷酸接点的非编码链,或一段与5′末端标记的与MMTV转录单位+28~+52碱基互补的寡核苷酸链杂交。利用T4多核苷酸激酶和[γ-32P]-ATP[111 GBq(3Ci)/mmol,NEN-Dupont公司]标记MMTV及β-肌动蛋白探针,然后洗膜,使用分子动态磷光计数仪检测杂交信号。
三、体外连续转录
从Ad5/MA和Ad5/MtplA感染的293细胞中分离细胞核成分,并将其悬于含有体积分数为40%的甘油、5 mmol/L MgCl2和0.1 mmol/L EDTA的50 mmol/L的Tris-HCl(pH8.3)中,计数浓度为108个核/ml。在30 ℃下延长起始RNA 30分钟,纯化标记RNA。标记的RNA与结合在尼龙膜上的Spel接头的寡核苷酸(pH 2.3)和PS73-ML质粒DNA杂交。杂交后冲洗、放射自显影方法同前。用闪烁计数方法测定膜相应部位杂交到不同DNA上的RNA数量。
, 百拇医药
四、β-肌动蛋白微基因转录本5′末端作图
由图1可见MA与MtplA基因核内和胞浆中5′端转录本。通过引物延伸法做图,该引物为与MMTV转录单位+28~+52位互补的片段。
五、加工后β-肌动蛋白微基因转录本的分析
分离纯化Ad5/MA或Ad5/MtplA感染细胞的胞浆和胞核RNA,以及分离含Poly(A)和不含Poly(A)成分的步骤如前所述。用逆转录聚合酶链反应(RT-PCR)法分离RNA,得到的扩增产物为杂交到人β-肌动蛋白微基因上游Poly(A)位或与Ad5基因4014-4055结合的引物延伸出来的片段。后者的序列位于重组病毒β-肌动蛋白小基因下游的E1B基因中(图1)。逆转录反应体系里含有200 U/μl的逆转录酶。PCR反应体系为:2.5 U的Taq聚合酶,2 mmol/L MgCl2,100 μmol/L dNTPs或100 μmol/L dATP、dGTP和dTTP,以及1 μmol/L dCTP、370 kBq(10 μCi)[α-
, 百拇医药
32 p]-dCTP(NEN-Dupont公司),与MMTV转录单位+21~+44位互补的3′引物,与3′端ML三联先导序列互补的5′引物和3′β-肌动蛋白引物。60个循环后,酚-氯仿抽提PCR产物,2倍体积乙醇沉淀,然后在1%琼脂糖凝胶上电泳,电泳缓冲液为TAE,用溴化乙淀染色或待凝胶干燥后,用放射自显影测定。
Ad5/MA和Ad5/Mtpl A的区别为后者在MMTV启动子与β-肌动蛋白基因间有一段主要晚期三联先导序列的导入,折线箭头为MMTV LTR启动子未剪割RNAs的启始,虚线表示人β-肌动蛋白基因的mRNA图1 激素诱导型β-肌动蛋白报导基因病毒
重组结构示意图
结果
一、Northern杂交检测报告基因产物
, 百拇医药
分离感染晚期细胞核及细胞浆RNA比较试验发现,带有tpl顺序的报告RNA更有效地聚积到胞浆,这可能是RNA加工、转送至胞浆的速率不同的结果。Northern杂交测定经地塞米松处理由MA和MtplA感染细胞的RNA,用插入到β-肌动蛋白微基因的寡核苷酸作分子探针,杂交结果显示一条与MA和mtplA报告基因剪接转录物的长度相吻合的条带,而且两种病毒报告基因RNA在胞浆中的分布不同。胞浆中的MA RNA是MtplA RNA的1/3,与此相反,MA RNA在胞核中的浓度明显大于MtplA RNA在胞核中分布。腺病毒报告基因特异性寡核苷酸及MMTV探针杂交显示的结果一致(图2,3)。
来自于Ads/MA感染Poly(A)细胞浆RNA及细胞核RNA(N)(3,5号样),Ad5/Mtp1A(2,4号样)感染细胞RNA,野生型Ad5(6号样)感染细胞RNA,与报告基因插入特异性的Spel寡核苷酸杂交图2 表达基因RNA杂交分析
, 百拇医药
图3 分子动力磷构象分析(Molecular Dynamics Phosphorimager)
测定分离的杂交片段,得出两组报告基因感染细胞后
细胞浆及细胞核RNA分布
二、脉冲示踪标记法
为了更精确地评估MA和MtplA RNA在胞浆中的稳定性对胞质浓度差异的作用,采用脉冲示踪标记法检测新合成的报告NRA在核/浆中的比例。结果发现,感染晚期,[3H]-Uridin标记经地塞米松处理的293细胞,在标记15分钟,示踪20分钟,与其相关RNA杂交后进行报告RNA、Hsp70和L2病毒RNA浓度分布的比较,新合成的Hsp RNA和L2病毒RNA在核/浆分布与预料相同:病毒晚期mRNA高效地进入胞浆,而Hsp70 RNA的输出受抑制。细胞质中观察到新合成带有tpl序列的报告基因RNA浓度大于不含tpl序列报告的RNA浓度。而细胞核中这两种报告RNA的浓度刚好相反,它们的RNA总浓度相同。带tpl序列的RNA进入胞浆增加表明转运速率高。用短时间脉冲示踪可有效地排除因时间和实验温度、条件所引起的数据误差,并证明带有tpl序列的报告RNA进入胞浆的效率是缺失tpl序列RNA的2.4~4.1倍(附表,图4~8)。
, 百拇医药
在同样的293细胞密度上用30 pfu/细胞的两种病毒(Ad/MA,Ad/Mtp1A)感染细胞,10小时后经地塞米松诱导,设感染对照(M),诱导(1+),无诱导(1-),14小时后,收集细胞核提取物作体外继续转录试验(图4).另一份感染细胞经[3H]-尿嘧啶进行示踪标记(图5)图4、5 β-肌动蛋白报告基因经诱导后在Ad/MA
和Ad/Mtp1A感染293细胞中的表达
三、腺病毒报告基因RNA的加工
引物延伸两种报告基因,在MMTV转录启始点+27~+52合成互补寡核苷酸片段,引物扩增的两种重组病毒都产生52个核苷酸的cDNA。Northern杂交和RT-PCR都没有发现带有病毒E1B序列的重组病毒特异性RNA,证实MA和MtplA的转录是独立系统,两种报告基因的转录启始不受残留的E1A增强子影响(图2,3,9)。
, http://www.100md.com
附表 重组腺病毒感染细胞后新合成RNA的有效转送率(%) 组别
新合成的细胞浆/细胞核RNA
HSP70
ML-L2
β-actin
Reporter
1.对照细胞
3.1
ND
ND
Ad5/MA+Dex,11h
0.3
, 百拇医药
8.5
0.1
Ad5/MtplA+Dex,11h
0.3
8.2
0.5
2.对照细胞
3.0
ND
ND
Ad5/MA+Dex,11h
0.5
2.5
, http://www.100md.com
0.3
Ad5/MtplA+Dex,11h
-
2.3
1.0
3.对照细胞
9.7
ND
ND
Ad5/MA+Dex,11h
0.4
11.7
0.1
, 百拇医药
Ad5/Mtp1A+Dex,11h
0.4
12.4
0.4
注:1.新合成Hsp70、ML-L2和β-肌动蛋白报告RNA在细胞质与细胞核中分布的定量,数字经图6~8分析获得,2.ND=检测不到,-=没检测;Dex=地塞米松
图6 两种重组腺病毒感染细胞后细胞浆RNA分布
图7 两种重组腺病毒感染细胞后细胞核RNA分布
, 百拇医药
图8 两种重组腺病毒感染细胞后细胞核与细胞浆新合
成RNA分布
四、RT-PCR测定报告基因RNA长度
比较MA和MtplA RNA反转录产物的长度,发现MtplA的RNA比相应的MA RNA长150~170个碱基,恰好和预期的tpl序列长短(164)相吻合。并与剪接后去除内含子6的报告基因RNA的长度完全一致,这说明从报告转录物中剪接了内含子(图1,9)。
RNA经Ad5/MA和Ad5/mtplA感染,于地塞米松诱导后提取、纯化,应用人β-肌动蛋白4024~4055互补引物及5′+22/44引物.Ad5/MA RNA(4,8);Ad5/mtplA RNA(2,6)图9 RT-PCR测定腺病毒报告基因转录体
, http://www.100md.com
讨论
本研究利用腺病毒报告基因能被激素诱导等生物特性来研究腺病毒感染细胞后,其mRNA在转录后加工中,从胞核向胞浆的转运机制。报告基因的转录严格依赖于外源性激素,通过增强子上GRES激活MMTV转录调控区开始转录[1]。这证明报告基因的转录活性不受相邻腺病毒序列(如上游的E1A增强子)的影响。去除地塞米松后,报告基因可继续转录几小时。这是研究腺病毒感染细胞后转录的激活、转录物加工及核膜转运的理想模型。
在两株腺病毒感染细胞晚期,被地塞米松激活的报告基因转录物都是以剪接后产物形式进入细胞质,说明这些细胞浆中的MA和MtplA RNA是被特异性输出的。MA基因不带任何病毒序列,转录后外运到胞浆的量明显低于MtplA RNA,同时细胞本身的mRNA如β-肌动蛋白、热休克蛋白转送受抑制(图4,5,6,7,8,附表)。
主要晚期三联先导序列可能控制mRNA转录后多个代谢步骤,包括mRNA从胞核转送,胞浆中的稳定性调控及翻译过程[3]。tpl增强输出RNA的最直接机制可能是在感染细胞通过特异地识别某些病毒或细胞介导蛋白,以协助RNA进入核膜输出通道。tpl的作用可能类似于HIV-1 mRNA RRE。这个HIV-1 Rev可识别转录后剪接和未剪接的病毒RNA位点而协助向核外转运[4]。最可能与腺病毒tpl特异结合的蛋白是E1B-55 000和E4-34 000蛋白。然而,这些病毒早期蛋白允许选择输出无tpl序列的RNA和某些细胞mRNA,这提示它们不能特异地识别tpl“输出”信号,表明可能还有其它病毒或细胞蛋白参与tpl的输出功能。因为tpl是首次应用组装病毒方式发现其具有在细胞内转运RNA的能力的病毒序列,新发现的任何特异性识别核蛋白都会提供关于与RNA输出和调节分子机制有关的重要线索。
, 百拇医药
本文在文稿及图片的整理工作中得到吴静、尹芳、曹晓燕等的帮助,特此致谢
参考文献
1 Berkner KL, Sharp PA. Effect of tripartite leader on synthesis of a non-viral protein in an adenovirus 5 recombinant. Nuc Acids Res, 1985, 13:841-857.
2 Cullen BR, Malim MH. The HIV-1 Rev protein: prototype of a novel class of eukaryotic post-transcriptional regulators. TIBS, 1991, 16:346-350.
3 Derget SM, Moore C, Sharp PA. Spliced segments at the 5′ terminus of adenovirus 2 late mRNA. Proc Natl Acad Sci USA, 1997, 74:3171-3175.
4 Huang WL, Pruzan R, Flint SJ. In vitro transcription from the adenovirus E2 early promoter by RNA polymerase Ⅲ, Proc Natl Acad Sci USA, 1994, 91:1265-1269.
(收稿:1998-01-05 修回:1998-06-30), http://www.100md.com