�˲���߰ͨ��GATAת¼���Ӵ̼���ϵ��ϸ����ֳ��
���ߣ������� ���÷ �ⳬȺ B.H.Chong
��λ�������� ���÷ �㽭��ҽѧԺ����ҽԺ���ⳬȺ B.H.Chong �Ĵ�������������ʿ��ѧ��������ʿҽԺ
�ؼ��ʣ��˲���߰��CFU-Meg��GATA-2ת¼���ӣ���Ѫ��������
�ٴ�ѪҺѧ��־990201 ժҪ Ŀ�ģ��˽��˲���߰(GS)�Ծ�ϵ��ϸ��(CFU-Meg )�Ĵ̼���ֳ���ã��Ƚ�GS����������TPO��IL-3��GATAת¼���ص����յ����á������������˺�ITP���߹���CFU-Meg������GS�̼����顢��Ӿ���ƶ����� ����(EMSA)�����彺�ƶ�����(Supershift assay)�͵�������ӡ����(Western Blot)�������GS�̼�����CFU-Meg��ֳ���ʼ��������ķ�ʽ��ѪС����������(ITP)���ߵ�CFU-MegҲ��GS��Ⱦ��GS��TPO��IL-3�����յ�������������MO7eϸ������G ATA������DNA��ϵĻ��Լ�ʹGATA-2���������ӡ�GS+TPO��GS+IL-3��Эͬ���յ�GATA �����Խ�һ�����ߡ����ۣ�GS�ܹ�ͨ�����������Ӻ���ǿ������ �����ö��̼�CFU-Meg��ֳ��GATAת¼���ӣ���Ҫ��GATA-2������Ѫϸ����GS��Ӧ���ź� ����;����
, ��Ĵҽҩ
Ginsenosides Stimulate Proliferation of Human Mega-karyocytic Pr ogenitors Through GATA Transcription Factor
Gao Ruilan,Wu Chaoqun,Jin Jinmai et al
Affiliated Hospital,Zhejiang College of Traditional Chinese Medicine,Ha ngzhou,310006
Abstract Objective:To investigate the action of ginsenosides (GS) on megakaryocytic progenitors (CFU-Meg) and possible mechanisms.M ethods:Using bone marrow culture of CFU-Meg,electrophoretic mobility s hift assay (EMSA),Supershift assay and Western blotting analysis.Result s:GS promoted proliferation of CFU-Meg in a dose-dependent manner.CFU -Meg from patients with idiopathic thrombocytopenia purpura was response to GS. GATA binding activity and GATA-2 protein induced by GS was elevated in growth f actor-dependent MO7e cells.GATA binding activity induced by the combination of GS with thrombopoietin (TPO),or GS with interleukin-3(IL-3) were higher than t hose by GS,TPO,or IL-3 alone.Conclusions:GS may stimulate the proliferation of CFU-Meg by enhancing the effect of TPO or IL-3,and by its gr owth factor-like function,and GATA binding factors,mainly GATA-2,may mediate t he response of megakaryocytes to GS.
, ��Ĵҽҩ
Key words Ginsenosides CFU-Meg GATA transcription factor Haematopoietic growth factor
�˲���Ϊ��˥�ϡ�ǿ׳���Ͳ���Ѫҩ�������ٴ�����ǧ������ ʷ��Ŀǰ�����ⱨ�����˲���ȡ�������Ѫ�ܡ��ڷ��ڼ������������õ��о�������ѪҺѧ������о����� ��������ǰ�������˲���߰(GS)�ܹ��̼������˵������ϰ���ƶѪ���ߵĺ�ϵ����ϵ��ϸ�� ��ֳ��1��������ͨ���۲�GS��GATAת¼���ص����յ����ã��о�GS�Ծ�ϵ��ϸ ��(CFU-Meg)�Ĵ̼���ֳ���á�����GS�̼���Ѫ�������������������ӣ��ʲ�����������Ѫ С��������(TPO)������-3(IL-3)��������Ƚϡ�
1 ���Ϻͷ���
1.1 �˹���CFU-Meg���� ����Abgrall��2���ķ������Ը�������������걾ȡ������������г� ���߹ǣ�����������ķ�ѪҺ�����ߡ�������ϵ��5%ȥѪС�����AB��Ѫ�塢15%̥ţѪ�� ��1%BSA��10-5mol/L 2-ME��IL-3 20 ng/ml(R&D system)��TPO 30 ng/ml(Genetec h)��105�к�ϸ��/�ף���0.9%�ļ���ά����֧���ÿ�ݱ걾��3���ף��ú�5%CO 2�����ʪ�����������з���14 d������CFU-Meg����(>3��ϸ��)��ϴȥ����ά�أ��� ��ͿƬ����Ƭ������-��ķ��Ⱦɫ�۲��ϸ����̬����������CD41��CD42( ����ҽѧ��ѧԺ)��ǣ�APAAP������ɫʶ���ϸ����
, ��Ĵҽҩ
1.2 GS�̼����� ������GS�����ĩ�ɱ�Ժ��ҩ�о������ƣ�����Ϊ86.3%����IMDM����Һ�� �Ƴ�2 000 mg/L�����˳�����4�汣�档��ǰϡ�ͳɺ������������CFU-Meg��������Ũ�� Ϊ1��5��10��20��50��100 ��g/ml���Բ���GS��������Ϊ���գ�ʵ��ֳ���������(TPO��IL -3)���������������顣
1.3 ԭ����ѪС����������(ITP)���ߵ�CFU-Meg��GS�������� 20��ITP����Ϊ��Ժ�����סԺ���ˣ���ϰ��ڶ���ȫ��ѪҺѧ�������ⶨ�ı� ��3�����Լ����л�ѪҺѧ��ȫ��Ѫ˨��ֹѪѧ������������4������ 6����Ů14��������6��65��(��λ��34��)������ͿƬƽ����ϸ����152.9��19.8/ Ƭ��ITP���ߵĹ���CFU-Meg��������20 ��g/ml��GS���������飬��GS�����>30%Ϊ����C FU-Meg��GS���еı���
1.4 �˾�ϸ�����GS�����ͺ˵����Ʊ� ������������ϸ����MO7e��LK Ashman��ʿ����(Hanson�����о����� �Ĵ����� )������1�����Ծ�ϸ����Ѫ��Ӥ��������Ѫ�����������ں�15%̥ţѪ�塢10 ng/ml IL- 3��IMDM�������С���GS�̼�ǰ��ϸ��������������MO7eϸ��ȥ�������ӣ��ں�0.5%̥ ţѪ����18 h��Ȼ��ֱ����GS��TPO��GS+TPO��IL-3��GS+IL-3��ϸ����ҩ�����2 h�� �˵�����ȡ������Dinam�����ļ��Ը�����ϸ������PBSϴ�Ӻ������ڻ���ҺA�У������� ø���Ƽ�Leupeptin��antipain��aprotinin��pepstatin(Sigma)����Ϊ10 ��g/mlŨ�ȣ��� ������ϸ��Ĥ���ռ�ϸ���ˣ������ڻ���ҺC�У���������������ϸ���ˣ����ij���12 000 r/min��20 min��ȡ���壬�ⶨ����Ũ�ȡ�
, ��Ĵҽҩ
1.5 ��Ӿ���ƶ���������(Electrophoretic Mobility shift As say,EMSA)�Ϳ��彺�ƶ�����(Supershift Assay) ��10 ��g�˵�����20 ��l�ķ�ӦҺ�У���HEPES��NaCl��EDTA�����͡�BSA�� 2.5 ��g/ml poly(dI/dC)����4��������15 min��5����̽���þ�����GATA-1��G ATA-2��GATA-3ת¼���ӹ�ͬ���λ���˫���Ѻ˸��
5��-CAC TTG ATA ACA GAA AGT GAT AAC TC-3��
3��-TG AAC TAT TGT CTT TCA CTA TTG AGA-5��
̽�뾭ĩ�˱�Ƿ��æ�-32P ATP(Amersham)��Ǻ�cpmֵ��ϡ�ͺ�ȡ5 �� l(50 0 00 cpm)���뷴ӦҺ����˵�����30 min����Ӧ������6%�۱�ϩ���������Խ���Ӿ������ ��ռ��ȸɽ�����XƬ��������Ӱ�Դ�������������DNA��ϵ�GATAת¼���ӵĸ�����(GAT A binding complex)����ͬ��ʵ���ظ�3�顣
, ��Ĵҽҩ
1.6 Supershift Assay ������EMSA��ͬ����̽����˵����ǰ�ȼ���1.5 ��l(1 mg/ml)Trans CruzTMgel supershift���嵽20 ��l��Ӧ���Һ�У�4�����2 h��ʹ��ԭ�����Ϸ� Ӧ������Ϊ����GATA-1(N6)����GATA-2(C-20)�Ϳ�GATA-3(HG3-31)(������Santa Cruz )��Ȼ�����32P-ATP��ǵ�GATA�Ѻ�����̽��5 ��l������30 min��6%�۱�ϩ������ ���Խ���Ӿ������
1.7 ��������ӡ����������ȡ��(10��g����)����SDS������������Һ25��g���3min����Ʒ��SDS-PAGE����Ӿ��������������õ�ת�Ƶ�������ά��Ĥ�ϣ���1.2%BSA-TBS-T����Һ���Ĥ�Ϸ�������λ��������Կ���GATA-2(C-20)����0.5��g/ml�����½�Ϸ�Ӧ1h������ϴĤ����������������ø��ǵĶ���(1��2000ϡ��)��ECL�����(AMERSHAM)����1MIN���ع�1~10MIN����Ӱ���ù��ܶ�ɨ���Ƿ��������ͬ��ʵ���ظ�3�顣
, ��Ĵҽҩ
2 ���
2.1 GS������CFU-MEG�Ĵ̼���ֳ����
��ͼ1����������ϸ������14 d�ɼ����־�ϸ�����䡣����3��10��ϸ����� �ĵ�����ϸ�����䣻�ڶ�����ϸ����ϼ����еľ�ϸ���μ����䡣�����Ƿ���������� �ӣ�CFU-Meg��GS�������Գʼ���������ʽ����������������ʱ(TPO 30 ng/ml��IL-3 20 ng/ml)��GS 5��50 ��g/ml�ܹ���������20.1%��76.6%���������ڲ���GS�Ķ� ����(P��<0.01)��������������������ʱ����������CFU-Meg����2.1��0. 6/105ϸ����GS 5��50 ��g/ml���ܴ̼�����һ�������ļ���Ϊ5.2��1.3-11 .5��2.4/105ϸ������ʾGS 20 ��g/ml������̼�CFU-Meg��ֳ��ǡ��Ũ�ȡ�
2.2 ITP���ߵ�CFU-Meg��GS������ ��������20��ITP���ߵ�CFU-Meg����Ϊ21.3��2.4/105ϸ����GS 20 ��g/mlʱ������Ϊ30.8��3.2/105ϸ�������ڲ���GS�Ķ�����(P<0.001 )��ƽ��GS�����Ϊ33.3��4.1%��ITP���ߵ�CFU-Meg��GS�������Ը�����ͬ���� ����GS�����>30%��Ϊ��GS���еı�����20������12����CFU-Meg��ʾ��GS���У�GS�� ����Ϊ31.0%��75.5%��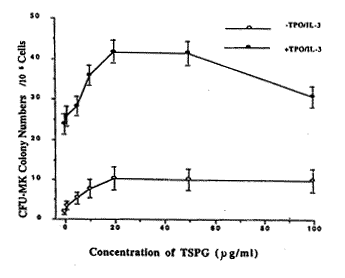
, ��Ĵҽҩ
ʵ���Ϊ���������Ӻ����������Ӷ���
ͼ1 ��ͬŨ�ȵ�GS��CFU-Meg�Ĵ̼���ֳ���� (n=9)
���� ITP���ߵ�CFU-Meg��GS�������� ����
�Ա�
����
�����
ϸ��/Ƭ
CFU-Meg/
105ϸ��
GS��
����(%)
, ��Ĵҽҩ
��GS
��GS
1
F
56
200
15.0
23.0
47.7
2
F
33
100
, ��Ĵҽҩ 5.3
6.0
11.0
3
M
44
35
26.3
42.6
38.5
4
F
48
50
, ��Ĵҽҩ
13.0
17.5
25.7
5
M
6
300
14.0
15.0
6.7
6
F
22
, http://www.100md.com 250
27.0
38.5
42.5
7
F
29
250
25.0
30.0
35.8
8
F
65
, http://www.100md.com
32
8.6
14.4
41.5
9
F
30
100
36.6
48.6
33.0
10
M
, ��Ĵҽҩ 34
250
29.0
40.0
36.0
11
F
48
80
22.6
38.3
69.4
12
, http://www.100md.com F
42
179
10.0
15.0
29.3
13
F
35
74
35.0
50.7
31.0
, ��Ĵҽҩ 14
M
8
200
22.0
29.0
17.5
15
F
34
117
17.3
29.3
21.0
, ��Ĵҽҩ
16
F
40
150
18.0
28.0
75.5
17
F
32
197
9.0
17.0
, http://www.100md.com 47.0
18
F
45
43
35.0
53.0
33.0
19
F
30
150
14.0
26.6
, ��Ĵҽҩ
12.8
20
M
8
300
43.8
53.6
11.0 ��s
��s
31.7
�� 21.5
21.3
��2.4
, http://www.100md.com
30.8
��3.2
33.3
��4.1
2.3 GS��TPO��GS��IL-3�յ�GATA������DNA��ϻ��� ��ͼ2��������������ϸ����MO7e��GS��TPO��IL-3�ֱ��̼�����2 h�� ��ȡ�˵�����EMSA�������ʾGS��TPO��IL-3�������Ƶ����ã�ϸ������GATA������DNA�� �ϻ������ӣ������������Դ��ܶ�����(Ӿ��2��3��5)����GS+TPO(Ӿ��4)����GS+IL-3(Ӿ ��6)ʱ��MO7eϸ�����ڵ�GATA������DNA��ϻ��Խ�һ�����ߣ������ڵ���GS��TPO��IL-3 ����ʾGS���������Ӷ��յ�GATA�����Ծ���Эͬ���á�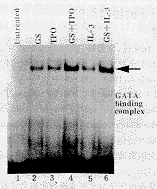
, ��Ĵҽҩ ͼ2 EMSA��ʾGS��TPO��IL-3�յ���GATAת¼ ���ص�����DNA��ϵĻ���
2.4 ����GS�յ���GATA������DNA��ϸ�����ijɷ� ��ͼ3���ӿ���GATA-1(Ӿ�� 3)��GATA-2(Ӿ�� 4)��GATA-3(Ӿ�� 5)������ �˵�����Ӧ����Supershift assay�������ʾGATA������DNA��������Ҫ��GATA-2����� �ϣ������������ʹ�����Դ������ƶ����ܶȼ��ͣ���ʾ����Ҫ�ɷ�ΪGATA-2ת¼���� ��
2.5 GS��TPO��IL-3����ϸ�������GATA-2������ ��ͼ4��MO7eϸ����GS+TPO(Ӿ�� 4)��GS+IL-3(Ӿ�� 6)�̼�������Western Bolt��ʾ�˵�����GATA-2�����Ϸ�Ӧ���������ܶ�ɨ�����GS+TPO��GS+IL-3������GAT A-2ת¼����������δ��������ϸ��������GS��TPO��IL-3������ʾGS���������ӵ�Эͬ�ԡ�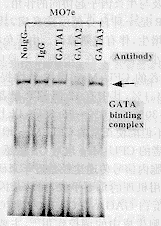
, http://www.100md.com
ͼ3 Supershift assay����GS�յ�GATA��DNA ���������Ҫ�ɷ�ΪGATA-2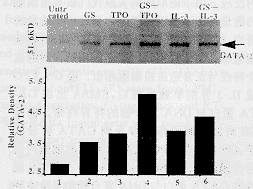
MO7eϸ����GS��TPO��IL-3�̼�����2 h��1Ϊδ��������2ΪGS��3ΪTP O��4ΪGS+TPO��5ΪIL-3��6ΪGS+IL-3
ͼ4 Western Blot����ϸ������GATA-2ת¼ ���ӵ�����
3 ����
������ϸ�������������ӵ����������£������ϲ����γ�CFU-Meg���䣬��GS 20 ��50 ��g/ml��ʹCFU-Meg�����2.1�ֱ���ߵ�11.6��11.5/105ϸ�������� ��������GS����ʱ����GS 5��50 ��g/ml�ܹ���������20.1%��76.6%�����ϸ ��ˮƽ˵��GS�̼�CFU-Meg��ֳ�����þ������������Ӽ�������������Эͬ�����á�
, ��Ĵҽҩ
ITP��һ�����������Լ�����������ϸ�����������г����ϰ������������ƣ�����Ѫ С����١���GS�̼������У�20��ITP���ߵ�CFU-Meg����ƽ�������Ϊ33.3��4.1 %��������12��ITP���ߵ�����ʴ���30%(31.0%��75.5%)�����Ͻ����ʾGS������ ʹ������CFU-Meg��ֳ������ITP���ߵ�CFU-MegҲ��GS���С�
ϸ�����źŴ���;���ǽ������о������������û�������Ҫ�ֶΣ���ת¼����GATA�嵰��GA GA-1��GATA-2��GATA-3��Ѫϸ�������еĵ���������7������Ҫ�ǽ鵼���������� �����ڷ�Ӧ����������ϸ����ֳ�ֻ����йػ���ı����ˣ����������о��������ӵķ� ����̽��GS�����û���������GATAת¼���ص��ײ�����Ѫϸ����GS�ķ�Ӧ��������ȡ������ ԭ����ϵ��ϸ��������ԶԶ��������ϸ�����źŴ���;�����о�����ѡ��������������MO 7eϸ���꣬��ϸ������������������Ӳ�����ֳ�������������������Ѫ��ϸ�����ơ��� GS��TPO��IL-3�ֱ�������EMSA����GATA������DNA������������Դ�����ʾGS��Ѫ ϸ���������漰��GATAת¼���ӣ�GS������TPO��IL-3���Ƶ�ϸ�����źŴ���;������Supe rshift assay����GS�յ���GATA������DNA������ijɷ���ҪΪGATA-2��ͬʱ��GATA-2�� ����Westernӡ����Ҳ������GS��TPO��IL-3���������ƣ����յ�����GATA-2������ ���ӡ����ױ���GATA-1���ϸ�������;�ϸ���ֻ��йأ�GATA-2���ػ�����������ӵ� ��Ӧ�漰��������Ѫϸ������ֳ��GATA-3��Ҫ�������ٺ�T�ܰ�ϸ����8����GATA-2 ������ﲻ�������ڵĺ�ϸ���������ڶ���DZ�ܵ���Ѫϸ�����ڼ��ĺ�ϵ��ϸ��������GATA -2�������ɴٽ�ϸ����ֳ������GATA-2����ȱ�ݵ�С����������ص�������Ѫ�����ϰ� ��9��10�������ĵĽ������Щ�о��������GSͨ��GATA-2ת¼���ӵ�������ֳ�й� �Ȼ��������ٽ���Ѫ��ϸ����ֳ��
, ��Ĵҽҩ
����GS���������ӵ�Эͬ���ÿ��ܱ�ֱ�ӵ����ø�Ϊ��Ҫ���˹���CFU-Meg�����γɵ�GS�� �����飬������������MO7eϸ����GS+TPO��GS+IL-3�̼���������GATAת¼���ص����� DNA��ϵĻ��ԣ��Լ�GATA-2���ĺ������߾����ڵ���GS��TPO��IL-3����Щ���֧���� ������裬GS�������������������⣬������ǿ��Ѫ�������ӵ�Эͬ���á�
�� Ϊ�й��ͰĴ�������������������Ŀ����Ĵ������������л�������
�������룺���ݣ�310006
�����
[1] ������������������÷����.�˲�����߰�������˺������ϰ���ƶ Ѫ������Ѫ��ϸ���Ĵ̼���ֳ����.�й�����ҽ�����־��1992��12(5)��258-287
, ��Ĵҽҩ [2] Abgrall J F,Berthou C,Sensebe L,et al.Decreased in vitro megakaryocyte colony formation in chronic idiopathic thrombocytopenic purpura.Br J Haematol,1993,85( 4):803
[3] �ڶ���ȫ��ѪҺѧѧ�������Ҫ.�л��ڿ���־��1984��33(11)��703
[4] ���л�ѪҺѧѧ��ȫ��Ѫ˨��ֹѪ�����йس�Ѫ����Ѫ��������ϱ�.�л�ѪҺѧ�� ־��1987��8(3)��183
[5] Wu Chaoqun,Ohmori Y,Bandyopadhyay S,et al.Interferon-stimulated response ele ment and NFKB sites cooperate to regulate double-Stranded RNA-induced transcri ption of IP-10 gene.J Interferon Res,1994,14:357
, http://www.100md.com
[6] Wu Chaoqun,Deng W,Ohmori Y,et al.Differential mechanisms of LPS-induc ed NFKB activation in macrophages and fibroblasts.J Endotoxin Res,1996,3:9
[7] Orkin SH.GATA-binding transcription factors in hematopoietic cells,Bl ood,1996,80(3):575
[8] Shivadasani RA,Orkin SH.The transcriptional control of hematopoiesis.Blood,19 96,87(10):4025
[9] Nagai T,Harigae H,Ishihara H,et al.Transcription factor GATA-2 is expressed in erythroid,early myeloid,and CD34+ human leukemia-derived cell lines.Bl ood,1994,84(4):1074
[10] Tsai FY,Keller G,Kuo FC,et al.An early haematopoietic defect i n mice lacking the transcription factor GATA-2.Nature,1994,371(5):221
( 1998-06-24 �ո�), http://www.100md.com
��λ�������� ���÷ �㽭��ҽѧԺ����ҽԺ���ⳬȺ B.H.Chong �Ĵ�������������ʿ��ѧ��������ʿҽԺ
�ؼ��ʣ��˲���߰��CFU-Meg��GATA-2ת¼���ӣ���Ѫ��������
�ٴ�ѪҺѧ��־990201 ժҪ Ŀ�ģ��˽��˲���߰(GS)�Ծ�ϵ��ϸ��(CFU-Meg )�Ĵ̼���ֳ���ã��Ƚ�GS����������TPO��IL-3��GATAת¼���ص����յ����á������������˺�ITP���߹���CFU-Meg������GS�̼����顢��Ӿ���ƶ����� ����(EMSA)�����彺�ƶ�����(Supershift assay)�͵�������ӡ����(Western Blot)�������GS�̼�����CFU-Meg��ֳ���ʼ��������ķ�ʽ��ѪС����������(ITP)���ߵ�CFU-MegҲ��GS��Ⱦ��GS��TPO��IL-3�����յ�������������MO7eϸ������G ATA������DNA��ϵĻ��Լ�ʹGATA-2���������ӡ�GS+TPO��GS+IL-3��Эͬ���յ�GATA �����Խ�һ�����ߡ����ۣ�GS�ܹ�ͨ�����������Ӻ���ǿ������ �����ö��̼�CFU-Meg��ֳ��GATAת¼���ӣ���Ҫ��GATA-2������Ѫϸ����GS��Ӧ���ź� ����;����
, ��Ĵҽҩ
Ginsenosides Stimulate Proliferation of Human Mega-karyocytic Pr ogenitors Through GATA Transcription Factor
Gao Ruilan,Wu Chaoqun,Jin Jinmai et al
Affiliated Hospital,Zhejiang College of Traditional Chinese Medicine,Ha ngzhou,310006
Abstract Objective:To investigate the action of ginsenosides (GS) on megakaryocytic progenitors (CFU-Meg) and possible mechanisms.M ethods:Using bone marrow culture of CFU-Meg,electrophoretic mobility s hift assay (EMSA),Supershift assay and Western blotting analysis.Result s:GS promoted proliferation of CFU-Meg in a dose-dependent manner.CFU -Meg from patients with idiopathic thrombocytopenia purpura was response to GS. GATA binding activity and GATA-2 protein induced by GS was elevated in growth f actor-dependent MO7e cells.GATA binding activity induced by the combination of GS with thrombopoietin (TPO),or GS with interleukin-3(IL-3) were higher than t hose by GS,TPO,or IL-3 alone.Conclusions:GS may stimulate the proliferation of CFU-Meg by enhancing the effect of TPO or IL-3,and by its gr owth factor-like function,and GATA binding factors,mainly GATA-2,may mediate t he response of megakaryocytes to GS.
, ��Ĵҽҩ
Key words Ginsenosides CFU-Meg GATA transcription factor Haematopoietic growth factor
�˲���Ϊ��˥�ϡ�ǿ׳���Ͳ���Ѫҩ�������ٴ�����ǧ������ ʷ��Ŀǰ�����ⱨ�����˲���ȡ�������Ѫ�ܡ��ڷ��ڼ������������õ��о�������ѪҺѧ������о����� ��������ǰ�������˲���߰(GS)�ܹ��̼������˵������ϰ���ƶѪ���ߵĺ�ϵ����ϵ��ϸ�� ��ֳ��1��������ͨ���۲�GS��GATAת¼���ص����յ����ã��о�GS�Ծ�ϵ��ϸ ��(CFU-Meg)�Ĵ̼���ֳ���á�����GS�̼���Ѫ�������������������ӣ��ʲ�����������Ѫ С��������(TPO)������-3(IL-3)��������Ƚϡ�
1 ���Ϻͷ���
1.1 �˹���CFU-Meg���� ����Abgrall��2���ķ������Ը�������������걾ȡ������������г� ���߹ǣ�����������ķ�ѪҺ�����ߡ�������ϵ��5%ȥѪС�����AB��Ѫ�塢15%̥ţѪ�� ��1%BSA��10-5mol/L 2-ME��IL-3 20 ng/ml(R&D system)��TPO 30 ng/ml(Genetec h)��105�к�ϸ��/�ף���0.9%�ļ���ά����֧���ÿ�ݱ걾��3���ף��ú�5%CO 2�����ʪ�����������з���14 d������CFU-Meg����(>3��ϸ��)��ϴȥ����ά�أ��� ��ͿƬ����Ƭ������-��ķ��Ⱦɫ�۲��ϸ����̬����������CD41��CD42( ����ҽѧ��ѧԺ)��ǣ�APAAP������ɫʶ���ϸ����
, ��Ĵҽҩ
1.2 GS�̼����� ������GS�����ĩ�ɱ�Ժ��ҩ�о������ƣ�����Ϊ86.3%����IMDM����Һ�� �Ƴ�2 000 mg/L�����˳�����4�汣�档��ǰϡ�ͳɺ������������CFU-Meg��������Ũ�� Ϊ1��5��10��20��50��100 ��g/ml���Բ���GS��������Ϊ���գ�ʵ��ֳ���������(TPO��IL -3)���������������顣
1.3 ԭ����ѪС����������(ITP)���ߵ�CFU-Meg��GS�������� 20��ITP����Ϊ��Ժ�����סԺ���ˣ���ϰ��ڶ���ȫ��ѪҺѧ�������ⶨ�ı� ��3�����Լ����л�ѪҺѧ��ȫ��Ѫ˨��ֹѪѧ������������4������ 6����Ů14��������6��65��(��λ��34��)������ͿƬƽ����ϸ����152.9��19.8/ Ƭ��ITP���ߵĹ���CFU-Meg��������20 ��g/ml��GS���������飬��GS�����>30%Ϊ����C FU-Meg��GS���еı���
1.4 �˾�ϸ�����GS�����ͺ˵����Ʊ� ������������ϸ����MO7e��LK Ashman��ʿ����(Hanson�����о����� �Ĵ����� )������1�����Ծ�ϸ����Ѫ��Ӥ��������Ѫ�����������ں�15%̥ţѪ�塢10 ng/ml IL- 3��IMDM�������С���GS�̼�ǰ��ϸ��������������MO7eϸ��ȥ�������ӣ��ں�0.5%̥ ţѪ����18 h��Ȼ��ֱ����GS��TPO��GS+TPO��IL-3��GS+IL-3��ϸ����ҩ�����2 h�� �˵�����ȡ������Dinam�����ļ��Ը�����ϸ������PBSϴ�Ӻ������ڻ���ҺA�У������� ø���Ƽ�Leupeptin��antipain��aprotinin��pepstatin(Sigma)����Ϊ10 ��g/mlŨ�ȣ��� ������ϸ��Ĥ���ռ�ϸ���ˣ������ڻ���ҺC�У���������������ϸ���ˣ����ij���12 000 r/min��20 min��ȡ���壬�ⶨ����Ũ�ȡ�
, ��Ĵҽҩ
1.5 ��Ӿ���ƶ���������(Electrophoretic Mobility shift As say,EMSA)�Ϳ��彺�ƶ�����(Supershift Assay) ��10 ��g�˵�����20 ��l�ķ�ӦҺ�У���HEPES��NaCl��EDTA�����͡�BSA�� 2.5 ��g/ml poly(dI/dC)����4��������15 min��5����̽���þ�����GATA-1��G ATA-2��GATA-3ת¼���ӹ�ͬ���λ���˫���Ѻ˸��
5��-CAC TTG ATA ACA GAA AGT GAT AAC TC-3��
3��-TG AAC TAT TGT CTT TCA CTA TTG AGA-5��
̽�뾭ĩ�˱�Ƿ��æ�-32P ATP(Amersham)��Ǻ�cpmֵ��ϡ�ͺ�ȡ5 �� l(50 0 00 cpm)���뷴ӦҺ����˵�����30 min����Ӧ������6%�۱�ϩ���������Խ���Ӿ������ ��ռ��ȸɽ�����XƬ��������Ӱ�Դ�������������DNA��ϵ�GATAת¼���ӵĸ�����(GAT A binding complex)����ͬ��ʵ���ظ�3�顣
, ��Ĵҽҩ
1.6 Supershift Assay ������EMSA��ͬ����̽����˵����ǰ�ȼ���1.5 ��l(1 mg/ml)Trans CruzTMgel supershift���嵽20 ��l��Ӧ���Һ�У�4�����2 h��ʹ��ԭ�����Ϸ� Ӧ������Ϊ����GATA-1(N6)����GATA-2(C-20)�Ϳ�GATA-3(HG3-31)(������Santa Cruz )��Ȼ�����32P-ATP��ǵ�GATA�Ѻ�����̽��5 ��l������30 min��6%�۱�ϩ������ ���Խ���Ӿ������
1.7 ��������ӡ����������ȡ��(10��g����)����SDS������������Һ25��g���3min����Ʒ��SDS-PAGE����Ӿ��������������õ�ת�Ƶ�������ά��Ĥ�ϣ���1.2%BSA-TBS-T����Һ���Ĥ�Ϸ�������λ��������Կ���GATA-2(C-20)����0.5��g/ml�����½�Ϸ�Ӧ1h������ϴĤ����������������ø��ǵĶ���(1��2000ϡ��)��ECL�����(AMERSHAM)����1MIN���ع�1~10MIN����Ӱ���ù��ܶ�ɨ���Ƿ��������ͬ��ʵ���ظ�3�顣
, ��Ĵҽҩ
2 ���
2.1 GS������CFU-MEG�Ĵ̼���ֳ����
��ͼ1����������ϸ������14 d�ɼ����־�ϸ�����䡣����3��10��ϸ����� �ĵ�����ϸ�����䣻�ڶ�����ϸ����ϼ����еľ�ϸ���μ����䡣�����Ƿ���������� �ӣ�CFU-Meg��GS�������Գʼ���������ʽ����������������ʱ(TPO 30 ng/ml��IL-3 20 ng/ml)��GS 5��50 ��g/ml�ܹ���������20.1%��76.6%���������ڲ���GS�Ķ� ����(P��<0.01)��������������������ʱ����������CFU-Meg����2.1��0. 6/105ϸ����GS 5��50 ��g/ml���ܴ̼�����һ�������ļ���Ϊ5.2��1.3-11 .5��2.4/105ϸ������ʾGS 20 ��g/ml������̼�CFU-Meg��ֳ��ǡ��Ũ�ȡ�
2.2 ITP���ߵ�CFU-Meg��GS������ ��������20��ITP���ߵ�CFU-Meg����Ϊ21.3��2.4/105ϸ����GS 20 ��g/mlʱ������Ϊ30.8��3.2/105ϸ�������ڲ���GS�Ķ�����(P<0.001 )��ƽ��GS�����Ϊ33.3��4.1%��ITP���ߵ�CFU-Meg��GS�������Ը�����ͬ���� ����GS�����>30%��Ϊ��GS���еı�����20������12����CFU-Meg��ʾ��GS���У�GS�� ����Ϊ31.0%��75.5%��
, ��Ĵҽҩ
ʵ���Ϊ���������Ӻ����������Ӷ���
ͼ1 ��ͬŨ�ȵ�GS��CFU-Meg�Ĵ̼���ֳ���� (n=9)
���� ITP���ߵ�CFU-Meg��GS�������� ����
�Ա�
����
�����
ϸ��/Ƭ
CFU-Meg/
105ϸ��
GS��
����(%)
, ��Ĵҽҩ
��GS
��GS
1
F
56
200
15.0
23.0
47.7
2
F
33
100
, ��Ĵҽҩ 5.3
6.0
11.0
3
M
44
35
26.3
42.6
38.5
4
F
48
50
, ��Ĵҽҩ
13.0
17.5
25.7
5
M
6
300
14.0
15.0
6.7
6
F
22
, http://www.100md.com 250
27.0
38.5
42.5
7
F
29
250
25.0
30.0
35.8
8
F
65
, http://www.100md.com
32
8.6
14.4
41.5
9
F
30
100
36.6
48.6
33.0
10
M
, ��Ĵҽҩ 34
250
29.0
40.0
36.0
11
F
48
80
22.6
38.3
69.4
12
, http://www.100md.com F
42
179
10.0
15.0
29.3
13
F
35
74
35.0
50.7
31.0
, ��Ĵҽҩ 14
M
8
200
22.0
29.0
17.5
15
F
34
117
17.3
29.3
21.0
, ��Ĵҽҩ
16
F
40
150
18.0
28.0
75.5
17
F
32
197
9.0
17.0
, http://www.100md.com 47.0
18
F
45
43
35.0
53.0
33.0
19
F
30
150
14.0
26.6
, ��Ĵҽҩ
12.8
20
M
8
300
43.8
53.6
11.0
��s31.7
�� 21.5
21.3
��2.4
, http://www.100md.com
30.8
��3.2
33.3
��4.1
2.3 GS��TPO��GS��IL-3�յ�GATA������DNA��ϻ��� ��ͼ2��������������ϸ����MO7e��GS��TPO��IL-3�ֱ��̼�����2 h�� ��ȡ�˵�����EMSA�������ʾGS��TPO��IL-3�������Ƶ����ã�ϸ������GATA������DNA�� �ϻ������ӣ������������Դ��ܶ�����(Ӿ��2��3��5)����GS+TPO(Ӿ��4)����GS+IL-3(Ӿ ��6)ʱ��MO7eϸ�����ڵ�GATA������DNA��ϻ��Խ�һ�����ߣ������ڵ���GS��TPO��IL-3 ����ʾGS���������Ӷ��յ�GATA�����Ծ���Эͬ���á�
, ��Ĵҽҩ ͼ2 EMSA��ʾGS��TPO��IL-3�յ���GATAת¼ ���ص�����DNA��ϵĻ���
2.4 ����GS�յ���GATA������DNA��ϸ�����ijɷ� ��ͼ3���ӿ���GATA-1(Ӿ�� 3)��GATA-2(Ӿ�� 4)��GATA-3(Ӿ�� 5)������ �˵�����Ӧ����Supershift assay�������ʾGATA������DNA��������Ҫ��GATA-2����� �ϣ������������ʹ�����Դ������ƶ����ܶȼ��ͣ���ʾ����Ҫ�ɷ�ΪGATA-2ת¼���� ��
2.5 GS��TPO��IL-3����ϸ�������GATA-2������ ��ͼ4��MO7eϸ����GS+TPO(Ӿ�� 4)��GS+IL-3(Ӿ�� 6)�̼�������Western Bolt��ʾ�˵�����GATA-2�����Ϸ�Ӧ���������ܶ�ɨ�����GS+TPO��GS+IL-3������GAT A-2ת¼����������δ��������ϸ��������GS��TPO��IL-3������ʾGS���������ӵ�Эͬ�ԡ�
, http://www.100md.com
ͼ3 Supershift assay����GS�յ�GATA��DNA ���������Ҫ�ɷ�ΪGATA-2
MO7eϸ����GS��TPO��IL-3�̼�����2 h��1Ϊδ��������2ΪGS��3ΪTP O��4ΪGS+TPO��5ΪIL-3��6ΪGS+IL-3
ͼ4 Western Blot����ϸ������GATA-2ת¼ ���ӵ�����
3 ����
������ϸ�������������ӵ����������£������ϲ����γ�CFU-Meg���䣬��GS 20 ��50 ��g/ml��ʹCFU-Meg�����2.1�ֱ���ߵ�11.6��11.5/105ϸ�������� ��������GS����ʱ����GS 5��50 ��g/ml�ܹ���������20.1%��76.6%�����ϸ ��ˮƽ˵��GS�̼�CFU-Meg��ֳ�����þ������������Ӽ�������������Эͬ�����á�
, ��Ĵҽҩ
ITP��һ�����������Լ�����������ϸ�����������г����ϰ������������ƣ�����Ѫ С����١���GS�̼������У�20��ITP���ߵ�CFU-Meg����ƽ�������Ϊ33.3��4.1 %��������12��ITP���ߵ�����ʴ���30%(31.0%��75.5%)�����Ͻ����ʾGS������ ʹ������CFU-Meg��ֳ������ITP���ߵ�CFU-MegҲ��GS���С�
ϸ�����źŴ���;���ǽ������о������������û�������Ҫ�ֶΣ���ת¼����GATA�嵰��GA GA-1��GATA-2��GATA-3��Ѫϸ�������еĵ���������7������Ҫ�ǽ鵼���������� �����ڷ�Ӧ����������ϸ����ֳ�ֻ����йػ���ı����ˣ����������о��������ӵķ� ����̽��GS�����û���������GATAת¼���ص��ײ�����Ѫϸ����GS�ķ�Ӧ��������ȡ������ ԭ����ϵ��ϸ��������ԶԶ��������ϸ�����źŴ���;�����о�����ѡ��������������MO 7eϸ���꣬��ϸ������������������Ӳ�����ֳ�������������������Ѫ��ϸ�����ơ��� GS��TPO��IL-3�ֱ�������EMSA����GATA������DNA������������Դ�����ʾGS��Ѫ ϸ���������漰��GATAת¼���ӣ�GS������TPO��IL-3���Ƶ�ϸ�����źŴ���;������Supe rshift assay����GS�յ���GATA������DNA������ijɷ���ҪΪGATA-2��ͬʱ��GATA-2�� ����Westernӡ����Ҳ������GS��TPO��IL-3���������ƣ����յ�����GATA-2������ ���ӡ����ױ���GATA-1���ϸ�������;�ϸ���ֻ��йأ�GATA-2���ػ�����������ӵ� ��Ӧ�漰��������Ѫϸ������ֳ��GATA-3��Ҫ�������ٺ�T�ܰ�ϸ����8����GATA-2 ������ﲻ�������ڵĺ�ϸ���������ڶ���DZ�ܵ���Ѫϸ�����ڼ��ĺ�ϵ��ϸ��������GATA -2�������ɴٽ�ϸ����ֳ������GATA-2����ȱ�ݵ�С����������ص�������Ѫ�����ϰ� ��9��10�������ĵĽ������Щ�о��������GSͨ��GATA-2ת¼���ӵ�������ֳ�й� �Ȼ��������ٽ���Ѫ��ϸ����ֳ��
, ��Ĵҽҩ
����GS���������ӵ�Эͬ���ÿ��ܱ�ֱ�ӵ����ø�Ϊ��Ҫ���˹���CFU-Meg�����γɵ�GS�� �����飬������������MO7eϸ����GS+TPO��GS+IL-3�̼���������GATAת¼���ص����� DNA��ϵĻ��ԣ��Լ�GATA-2���ĺ������߾����ڵ���GS��TPO��IL-3����Щ���֧���� ������裬GS�������������������⣬������ǿ��Ѫ�������ӵ�Эͬ���á�
�� Ϊ�й��ͰĴ�������������������Ŀ����Ĵ������������л�������
�������룺���ݣ�310006
�����
[1] ������������������÷����.�˲�����߰�������˺������ϰ���ƶ Ѫ������Ѫ��ϸ���Ĵ̼���ֳ����.�й�����ҽ�����־��1992��12(5)��258-287
, ��Ĵҽҩ [2] Abgrall J F,Berthou C,Sensebe L,et al.Decreased in vitro megakaryocyte colony formation in chronic idiopathic thrombocytopenic purpura.Br J Haematol,1993,85( 4):803
[3] �ڶ���ȫ��ѪҺѧѧ�������Ҫ.�л��ڿ���־��1984��33(11)��703
[4] ���л�ѪҺѧѧ��ȫ��Ѫ˨��ֹѪ�����йس�Ѫ����Ѫ��������ϱ�.�л�ѪҺѧ�� ־��1987��8(3)��183
[5] Wu Chaoqun,Ohmori Y,Bandyopadhyay S,et al.Interferon-stimulated response ele ment and NFKB sites cooperate to regulate double-Stranded RNA-induced transcri ption of IP-10 gene.J Interferon Res,1994,14:357
, http://www.100md.com
[6] Wu Chaoqun,Deng W,Ohmori Y,et al.Differential mechanisms of LPS-induc ed NFKB activation in macrophages and fibroblasts.J Endotoxin Res,1996,3:9
[7] Orkin SH.GATA-binding transcription factors in hematopoietic cells,Bl ood,1996,80(3):575
[8] Shivadasani RA,Orkin SH.The transcriptional control of hematopoiesis.Blood,19 96,87(10):4025
[9] Nagai T,Harigae H,Ishihara H,et al.Transcription factor GATA-2 is expressed in erythroid,early myeloid,and CD34+ human leukemia-derived cell lines.Bl ood,1994,84(4):1074
[10] Tsai FY,Keller G,Kuo FC,et al.An early haematopoietic defect i n mice lacking the transcription factor GATA-2.Nature,1994,371(5):221
( 1998-06-24 �ո�), http://www.100md.com