趋化因子及受体在T细胞致敏和Th1/Th2介导的免疫应答中的作用
作者:龚如军 杨俊伟
单位:杨俊伟 南京军区南京总医院 解放军肾脏病研究所(南京,210002);龚如军 南京大学医学院博士研究生
关键词:趋化因子;Th1/Th2细胞;免疫应答
T细胞在胸腺中生长发育成初始型T细胞
T细胞在胸腺中生长发育成初始型T细胞(naive T cell)后,通过循环流经全身血液及次级淋巴器官,搜寻抗原呈递细胞(antigen-presenting cell,APC)呈递的抗原。初始型T细胞发现其特异抗原后,即增殖分化为具有新的迁移能力的效应T细胞和记忆T细胞[1]。效应T细胞能够进入外周组织,识别抗原,产生炎性或变态反应;记忆T细胞除具有向外周组织迁移的能力外,还能迁移至淋巴结,进一步刺激产生新一批效应T细胞。因此,T细胞的功能与其迁移能力密切相关,其中必然存在特定的分子机制,使得相应的细胞能够迁移至相应的组织中去。
, 百拇医药
1 趋化因子及其受体是调控白细胞定位的重要成分[2]
白细胞向组织迁移时,在两个阶段受到调控。首先,白细胞在血管内皮表面的滞留及牢固粘附;其次,白细胞从内皮向间质并进一步向特定微环境的迁移。趋化因子在这两个过程中发挥了重要作用。白细胞与内皮的结合牵涉到选择素介导的滚动和整合素介导的牢固粘附。内皮细胞表面的趋化因子与其位于白细胞表面的相应受体相互作用触发了整合素的活化,导致牢固粘附。外周各种组织细胞生成的趋化因子驱使白细胞穿过组织移行至特定微环境。因此,选择素、趋化因子受体和整合素的不同表达为白细胞移行提供了一套“地址码”(addresscode),使白细胞能够在特定位置出现,定位于特定组织。
从上述过程不难得出如下假说:白细胞通过一种多步骤导航模式(multistep navigation mode),对一系列趋化因子先后产生反应。即借助一系列不同的趋化因子受体,使得白细胞能够完成由内皮经间质定位于目的微环境的过程。因此,有些细胞因子在滞留阶段发挥作用,有些则在后继各阶段起作用。显而易见,由于趋化因子及其受体的丰富多样性,对于不同的白细胞,趋化系统能够灵活准确地提供相应的迁移形式。
, 百拇医药
尽管,人们早就认识到选择素和整合素在初始型T细胞和效应/记忆T细胞上表达是不同的,但直到最近,才发现随着抗原刺激程度的不同以及分化程度的不一,许多趋化因子受体在T细胞上的表达也是不一样的。新近的研究表明,趋化因子与受体是调节T细胞及其伴随细胞定位、进一步致敏,以及调节Th1/Th2介导免疫应答所必需的。
2 趋化因子受体在T细胞上的表达
2.1 初始型T细胞和记忆T细胞趋化因子受体表达[3~8] T细胞趋化的早期研究中,人们发现只有记忆/活化T细胞对已知的趋化因子有反应,而初始型T细胞则无反应;而且,淋巴细胞在淋巴组织中的移行可以被百日咳毒素通过抑制趋化因子受体信号传递的作用而阻断。因此,早期研究仅认为只有记忆或活化T细胞才表达Burkitt淋巴瘤受体-1(BLR-1)、C-X-C亚族趋化因子受体5(CXCR5)及CXCR3等趋化因子受体,并对正常T细胞表达和分泌、活化时表达下降的因子(reduced upon activation,normal texpressed and secreted,RANTES)、单核细胞趋化蛋白-1(MCP-1)、-2、-3有反应。
, http://www.100md.com
近年来,人们对趋化因子的功能特性有了进一步认识。许多研究发现趋化因子大体可分为两大类:①炎症性趋化因子,外周组织炎症状态下可被诱导产生或被强烈上调表达;②构成性趋化因子,履行看家功能(housekeeping function),也可能参与构成性的白细胞移行。人类发现的第一个趋化因子即为炎症型,因此只有记忆/活化T细胞表达其同族受体,并对其有反应。最近,人们发现初始型T细胞上也表达趋化因子受体,包括CXCR4、C-C亚型趋化因子受体7(CCR7)。此外,初始型T细胞还表达一种与树突状细胞趋化因子-1(DC-CK1)作用有关的受体,但其结构和功能目前尚不清楚。
T细胞表面趋化因子受体从构成型向炎症型的转变,是T细胞活化、分化的结果,同时它也将改变T细胞的迁移习性。
2.2 Th1、Th2细胞趋化因子受体的表达 与初始型T细胞不同,抗原刺激过的T细胞是不均一的,从另一方面也反应了T细胞活化状态和分化程度的不同。T细胞应答的中心过程是T细胞向Th1型或Th2型分化[9]。T细胞致敏阶段细胞因子的不同组合决定了效应T细胞所产生的细胞因子的类型。CD4+Th1能够产生γ干扰素(IFN-γ),活化单核-巨噬细胞,因此可以杀伤细胞内微生物。相反,CD4+Th2细胞则产生IL-4和IL-5,并与IgE、嗜酸和嗜碱粒细胞主导的反应相关。在MHC-I限制的CD8+T细胞应答中也观察到Th1、Th2的分化。作为Th1、Th2不同效应能力的一部分,它们可能表达不同的趋化因子受体,使得它们向不同组织趋化。
, http://www.100md.com
2.2.1 Th2细胞特异表达的趋化因子受体[10~11] CCR3是嗜酸粒细胞趋化因子(eotaxin)、eotaxin2、RANTES、MCP-2,-3,-4的受体,最初在嗜酸/嗜碱细胞中发现。Th2细胞上选择性地表达CCR3。使用CCR3特异的单克隆抗体检测发现,成人外周血小部分记忆/效应T细胞有CCR3分布,这些细胞同时表现为CD45R0-;分选出这些细胞,体外扩增后发现能产生高浓度的IL-4,IL-5,从而证明这些细胞大多为Th2细胞。体外分化的Th2细胞系也表达CCR3,但Th1则未见表达。此外,不论CD4+还是CD8+T细胞群,CCR3+细胞组分与生成IL-4的细胞数均相近。因此,CCR3+的T细胞可以产生IL-4,而生成IL-4是Th2细胞的重要特征。
目前还不清楚CCR3是否是决定Th2细胞迁移的主要趋化因子受体。除CCR3外,Th2细胞还表达CCR4(TARC的受体),并且在加入胸腺和活化调节的趋化因子(TARC)后,分离出能够动员Ca2+的T细胞,发现这些细胞也能产生IL-4。但CCR4并非Th2的特异标记,它的表达有时可能和产生IL-4的表型相分离;如有文献报道在转化生长因子β(TGF-β)作用下,T细胞系虽然能表达CCR4但并不产生IL-4或IFN-γ。因此只有CCR3才始终与Th2产生IL-4的表型相一致。
, 百拇医药
2.2.2 Th1细胞特异表达的趋化因子受体[11~14] 与Th2细胞能够选择性地表达趋化因子受体一样,Th1也选择性地表达某些受体。CXCR3就是其中之一,它是趋化因子IP-10(IFN-γ诱导蛋白-10)、Mig(IFN-γ诱导的单核因子)和I-TAC(IFN-γ可诱导的T细胞α趋化诱导剂)的受体,所有这些趋化因子均可被IFN-γ上调。CXCR3在大多数外周血记忆T细胞都有表达,但Th1表达水平远高于Th2,与此相应,Th1对IP-10反应所需的剂量仅为Th2的十分之一。
CCR5是RANTES、巨噬细胞炎性蛋白-1α(MIP-1α)和MIP-1β的受体,但是否为Th1的特异性受体尚有争议。虽然有研究发现CCR5选择性地表达于Th1,但Sallusto等的工作证实,尽管CCR5在Th1上有表达,而Th2没有表达,但在新近活化的T细胞,不管其功能、分化如何都有同等量的CCR5表达。使用CCR5特异性抗体,发现CCR5存在于一小部分CD26+、L-选择素(L-selectin)、CD45R0+的外周血T细胞,而这一系列表型均与T细胞的新近活化相一致。此外,当撤减IL-2时,CCR5则迅速消失,而CCR3、CXCR3却无变化。因此CCR5+T细胞可能仅代表最新产生的循环效应细胞。
, http://www.100md.com
尽管现有资料十分有限,但已有的研究发现Th1、Th2表达不同的趋化因子受体,因而对趋化因子反应也不相同。近期对小鼠淋巴系统的研究也已观察到类似现象。
3 IFN-α和TGF- β调节趋化因子受体的表达[11,15~17]
T细胞的分化不仅受抗原刺激强度的调节,同时还受致敏阶段周围细胞因子的影响。这些促进T细胞分化的细胞因子来源于APC、应答的T细胞或周围旁观细胞。尽管IL-2和IL-4在Th1、Th2分化过程中起着主要作用,但其它细胞因子同样也影响T细胞的最终分化。即便在没有IL-12或仅存在IL-4的环境下,IFN-α仍可诱导人Th1分化。而TGF-β则能够抑制人T细胞分化,使增殖的T细胞处于半初始化(semi-naive)状态。半初始化T细胞除IL-2外,不生成其它任何细胞因子,但在适当刺激后,仍可进一步分化为Th1或Th2。
Sallusto等研究发现,IFN-α和TGF-β除影响分化外,还能显著地影响不同类型趋化因子受体的表达。在Th1或Th2分化环境中加入IFN-α后,CCR3、CCR4的表达受到抑制,而CCR1的表达上调。因此,与IL-12诱导分化的Th1相比,IFN-α诱导分化的Th1细胞还上调表达有大量的CCR1,而CCR1能使T细胞对更多的趋化因子作出反应。相反,TGF-β抑制CCR2、CCR3和CCR5的表达,而上调CCR4、CCR7的表达。所以,与其抑制分化的作用相一致,那些能促进T细胞向外周组织迁移的趋化因子受体也受到TGF-β的抑制;而有助于T细胞归巢的趋化因子受体则得到增强表达。总之,IFN-α、TGF-β能够调节T细胞趋化因子受体的表达,灵活地调节着白细胞的运动,对特定组织中细胞的组成也有深刻的影响(见图1)。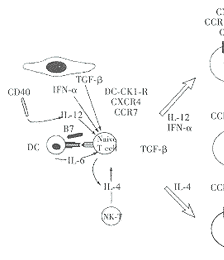

, http://www.100md.com
图1 T细胞趋化因子受体表达的调节
DC:树突状细胞 DC-DK1-R:树突状细胞趋化因子1受体
Naive T cell:初始型T细胞 DTH:迟发型超敏反应
4 趋化因子协调次级淋巴器官中T细胞、B细胞和树突状细胞的关系[18]
趋化因子及其受体在引导特定细胞迁移至特定炎症组织及淋巴器官,诱发初级免疫应答产生的过程中也发挥了重要作用(见图2)。
淋巴结中初始型T细胞和树突状细胞(DC)充分相互作用,是T细胞致敏的重要场所。未成熟的DC及单核细胞(DC的直接前体),表达多种受体,如CCR1、2、5、6,迅速募集到外周炎症组织。进入炎症组织后,DC立即俘获抗原,同时受炎症因子,如TNF-α、IL-1或细菌病毒产物(如LPS或dsRNA等)的刺激而分化成熟。DC的成熟不仅出现粘附分子或协同刺激分子上调,而且可使之向次级淋巴器官迁移。LPS或TNF-刺激DC产生大量炎症型趋化因子,进一步募集未成熟的DC;并可使其自身的趋化因子受体下调,有利于DC及时从炎症组织中退出。当DC的炎症型趋化因子受体下调时,构成性趋化因子受体如CCR4、CCR7、CXCR4在成熟的DC得到上调表达。CCR7对引导成熟的DC进入淋巴管尤为重要,这是因为淋巴管内皮细胞能够产生CCR7的同族配体——次级淋巴组织趋化因子(SLC)。DC从淋巴管引流到局部淋巴结,淋巴结中固有的成熟DC可产生另一种CCR7配体EBI1配体趋化因子(ELC),控制DC定位于淋巴结的T细胞区。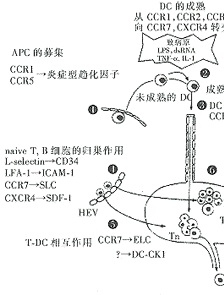
, http://www.100md.com
图2 趋化因子及其受体在T细胞致敏和Th1/Th2介导的免疫应答中的作用
APC:抗原呈递细胞 naive T:初始型T细胞 SLC:次级淋巴样组织趋化因子
DC:树突状细胞 Mig:γ-干扰素诱导的单核因子 ELC:EBI1配体趋化因子
MIP:巨噬细胞炎性蛋白DCA-1:B细胞趋化因子1MDC:巨噬细胞来源的趋化因子
SDF-1:基质细胞来源的因子1PSGL-1:P-selectin糖蛋白配体1IP-10:干扰素可诱导蛋白10
与DC相伴的初始型T细胞,通过另一条途径——高内皮细胞的毛细血管后微静脉(HEV)进入淋巴结。L-selectin和淋巴细胞功能相关抗原-1(LFA-1)在该过程中起促进作用。淋巴组织中至少产生包括基质细胞来源的因子-1(SDF-1)、SLC、ELC在内的三种趋化因子,可以使血循环中T细胞迅速滞留于细胞间粘附分子-1(ICAM-1)表达区域,而它们相应的受体CXCR4和CCR7在初始型T细胞均呈高水平表达。因此,CXCR4和CCR7同时也可能是介导牢固粘附和穿过HEV迁移的重要趋化因子受体。初始型T细胞进入淋巴结后,自由地渗滤通过没有细胞外基质的副皮质区,使之与DC充分作用。DC自身也分泌ELC或DC-CK1等趋化因子,除了对初始型T细胞有活性,吸引T细胞外,还可能在T细胞致敏的起始步骤中起作用。
, 百拇医药
初始型B细胞,与初始型T细胞一样,也表达CXCR4和CCR7,从HEV进入淋巴结后定位于滤泡B细胞区。初始型B细胞还表达CXCR5,通过基因打靶技术业已证实CXCR5是淋巴滤泡及生发中心形成所必需的成份。人类CXCR5的配体是B细胞趋化因子-1(BCA-1),小鼠中的相应物质为B淋巴细胞趋化剂(BLC),它们均由滤泡中的细胞产生,在吸引初始型B细胞以及活化的Th细胞、形成滤泡结构的过程中发挥作用。活化的B细胞也能生成巨噬细胞衍生的趋化因子(MDC)等趋化因子,可能促进T细胞和B细胞的相互作用。
5 趋化因子影响白细胞迁移的作用
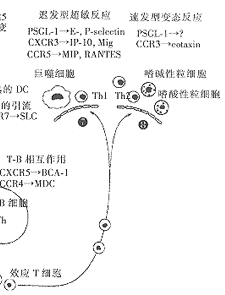
5.1 迟发型超敏反应和变态反应中白细胞迁移的调节[11,13,19] 局部组织的炎症增强了白细胞内皮的粘附和移行。这一现象的基础是内皮细胞粘附分子的上调以及内皮细胞和组织细胞分泌多种炎症型趋化因子,这些物质分布的差异有可能决定哪些细胞将会优先聚集到某一特定的炎症部位(见图2)。
, http://www.100md.com
在迟发型超敏反应的组织中,炎症性细胞因子,如TNF-α、IL-1和IFN-γ,能够促进内皮细胞表达E-selectin、P-selectin和血管细胞粘附分子-1(VCAM-1),并刺激产生多种趋化因子,包括IP-10、Mig。Th1表达P-selectin糖蛋白配体-1(PSGL-1),可与E-selectin、P-selectin结合,涉及T细胞移行到迟发型超敏反应区。Th2上虽然也曾发现PSGL-1,但它并不与E-selectin、P-selectin结合,有研究显示,它可能与变态反应组织内皮上优先表达的某种未知配体功能相关。
最近的研究表明,有些趋化因子受体并不依赖于整合素的作用,是直接在粘附过程中发挥作用的。NK和一些活化T细胞上均表达CX3CR1,因此可与fractalkine(内皮细胞上一种整合性膜趋化因子)相作用而介导滚动、粘附。CCR1也有类似作用,它可将E-selectin、P-selectin作为替代配体与之结合。此外有研究发现,除LFA-1外,记忆/活化T细胞还表达很晚出现的抗原-4(VLA-4)和其它整合素;虽然VLA-4与其配体VCAM-1的结合不需要趋化因子的参与,但是,VLA-4与纤维连接蛋白的作用却必须要有趋化因子参与,而且纤维连接蛋白在T细胞移行到组织中起了重要作用。尽管人们目前尚不清楚IP-10和Mig,哪一个在粘附或移行水平发挥作用,但由于它们的表达是受调节的,且其同族受体CXCR3在Th1上呈高水平的表达,因此这些趋化因子可能在选择性地募集Th1至以IFN-γ为主的炎症部位起重要作用。此外,由于T细胞均表达高水平的炎症性趋化因子相应的同族受体,如CCR5,因此RANTES、MIP-1α和MIP-1β还可以吸引新近活化的T细胞。
, 百拇医药
与变态反应关系最为密切的趋化因子为嗜酸性粒细胞趋化因子(eotaxin),它在肺中生成,并在肺中保持高浓度。eotaxin仅与CCR3结合,而Th2上选择性地表达CCR3。因此,eotaxin可以募集Th2及其它炎症细胞到变态反应组织中。除eotaxin外,其它趋化因子也可能参与了变态反应,在变态反应不同阶段,募集不同的效应细胞。
5.2 趋化因子受体在白细胞共定位中的作用[14] 我们常可以通过一个T细胞的伴随细胞来判断该T细胞的种类。Th2常伴随有嗜酸和嗜碱性细胞,而Th1则更常与巨噬细胞和中性粒细胞共处。这可能是由于抗原特异性T细胞及其它不同类型的效应细胞共享相同的趋化因子受体,而使它们共处一处。例如,Th2、嗜酸、嗜碱细胞均普遍表达CCR3。这三种细胞能够协同作用产生变态炎症反应,但更重要的是嗜酸、嗜碱细胞的活化、存活依赖于Th2分泌的IL-5,IL-4。再如,Th1细胞和单核细胞均普遍表达CCR1和CCR5,单核细胞是巨噬细胞和DC的前体。这就使得APC、清道夫细胞和炎症T细胞能同时募集到迟发型超敏反应组织中。
, 百拇医药
对炎症组织中T细胞的趋化因子受体的直接分析已证实了上述理论。因此,在变态反应,如鼻息肉或接触性皮炎都有CCR3+的T细胞、大量嗜酸/嗜碱性粒细胞浸润;而迟发型超敏反应,如类风湿关节炎,则仅有CCR5+和CXCR3+的T细胞浸润。类风湿关节炎时,滑膜液中有大量的CCR5+T细胞,而血液及淋巴组织中的数量则相对很少,这从另一方面说明T细胞定位是有选择的。
NK细胞是先天免疫的重要效应细胞,目前对NK在体内移行的途径知之甚少。NK细胞表达多种趋化因子受体,能使其和其它白细胞一起成为机体的首道防线。已有研究报道,NK可对多种趋化因子均有反应,包括IP-10,LT(lymphotactin)、RANTES、MIP-1、MCP-1,2,3、MDC。
结语[20] 大量趋化因子及其受体的发现,使我们进一步认识到淋巴细胞是如何选择性地移行至组织的。目前,我们面临的首要任务是澄清所有趋化因子受体的功能和表达形式。各种受体在不同细胞中的信号传递能力也是一个重要的研究领域,但目前仍知之甚微。一些受体可能采用不同的信号途径,因此,不仅在移行,在淋巴细胞的活化、转化甚至存活中都可能发挥作用。此外,趋化因子受体信号传递能力不同,也可能是由于G蛋白信号系统的调节蛋白表达不同所致;这些蛋白的表达对决定趋化因子受体的功能至关重要。另外一个重要的研究领域是组织中趋化因子生成的调控,现已清楚,免疫反应的性质是由组织中生成的趋化因子的种类所决定的。
, http://www.100md.com
趋化因子及其受体可能是当今治疗炎症疾病最有希望的研究领域。多步骤导航模式,即一个迁移细胞在其移行的不同特定阶段可能表达不同的趋化因子受体、对不同的趋化因子有反应;因此,如果阻断趋化因子受体则会导致迁移细胞迷失方向,不能到达其最终目标。该理论同时也解释了为何阻断人类趋化因子可以非常有效地抑制多种炎症反应。趋化因子的功能是使白细胞选择性地向特定组织移行,所以拮抗趋化因子受体的治疗将比现有疗法更具选择性,产生更少的免疫抑制副作用。目前,已生产的趋化因子受体拮抗剂多种多样,包括单克隆抗体,趋化因子修饰物以及最近研制的小分子无机物。在未来的几年中,它们很可能成为治疗自身免疫疾病和变态反应疾病的有效药物。
总之,趋化因子已从未知的细胞因子的分枝学科发展成为一个涉及生物学许多方面的新研究领域。趋化因子及其受体在生长发育、上皮和内皮细胞移行、神经生物学、T细胞活化和感染性疾病中的作用正逐步为人类所认识。
参考文献
, 百拇医药
1 Mackay CR.Immunological memory.A dv Immunol,1993,53:217
2 Picker LJ,Butcher EC.Physiological and molecular mechanisms of lymphocyte homing.Annu Rev Immunol,1992,10:561
3 Bargatze RF,Butcher EC.Rapid G protein -regulated activation event involved in lymphocyte binding to high endothelial v enules.J Exp Med,1993,178:367
4 Foster R,Emrich T,Kremmer E et al.Ex pression of the G-protein-coupled recept or BLR1 defines mature,recirculating B cells and a subset of T-helper memory cel ls.Blood,1994,84:830
, 百拇医药
5 Loetscher P,Seitz M,Baggiolini M et al.Interleukin-2 regulates CC chemokine receptor expression and chemotactic resp onsiveness in T lymphocytes [see comment s].J Exp Med,1996,184:569
6 Schall TJ,Bacon K,Toy KJ et al.Selec tive attraction of monocytes and T lymph ocytes of the memory phenotype by cytoki ne RANTES.Nature,1990,347:669
7 Yoshida R.Imai Y,Hieshima K et al.Mo lecular cloning of a novel human CC chem okine EBI1-ligand chemokine that is a specific functional ligand for EBI1,CCR7.J Biol Chem,1997,272:13803
, 百拇医药
8 Hieshima K,Imai T,Baba M et al.A novel human CC chemokine PARC that is most homologous to macrophage-inflammatory protein-1 alpha/LD78 alpha and chemotactic for T lymphocytes,but not for monocytes .J Immunol,1997,159:1140
9 杨俊伟,黎磊石.Th1/Th2细胞与免疫.肾脏病与透析肾移植杂志,1995,4:533
10 Ponath PD,Qin S,Post TW et al. Molecular cloning and characterization of a human eotaxin receptor expressed sel ectively on eosinophils [see comments].J Exp Med,1996,183:2437
, http://www.100md.com
11 Sallusto F,Lenig D,Mackay CR et al.Flexible programs of chemokine rece ptor expression on human polarized T hep er 1 and 2 lymphocytes.J Exp Med,1998,18 7:875
12 Luster AD,Ravetch JV.Biochemical characterization of a gamma interferon- inducible cytokine(IP-10).J Exp Med,1987 ,166:1084
13 Bonecchi R,Bianchi G,Bordignon P P et al.Differential expression of che mokine receptors and chemotactic respons iveness of type 1 T helper cells(Thls)and Th2s.J Exp Med,1998,187:129
, 百拇医药
14 Loetscher P,Uguccioni M,Bordoli L et al.CCR5 is characteristic of Th1 lymphocytes [letter].Nature,1998,391:344
15 刘俊铎,刘志红.Th1/Th2细胞与自身免疫性疾病.肾脏病与透析肾移植杂志,1997,6:555
16 Cella M,Scheidegger D,Palmer Leh mann K et al.Ligation of CD40 on dendr itic cells triggers production of high l e vels of interleukin-12 and endhances T cell stimulatory capacity:T-T help via A PC activation.J Exp Med,1996,184:747
, 百拇医药
17 Rogge L,Barberis-Maino L,Biffi M et al.Selective expression of an inte rleukin-12 receptor component by human T helper 1 cells.J Exp Med,1997,185:825
18 Sozzani S,Sallusto F,Luini W et al.Migration of dendritic cells in res ponse to formyl peptides,C5a,and a disti nct set of chemokines.J Immunol,1995,155 :3292
19 Taub DD,Lloyd AR,Conlon K et al .Recombinant human interferon-inducible protein 10 is a chemoattractant for hum an monocytes and T lymphocytes and promo tes T cell adhesion to endothelial cells .J Exp Med,1993,177:1809
20 Strieter RM,Standiford TJ,Huffin agle GB et al.“The good,the bad,and the ugly.”The role of chemokines in models of human disease.J Immunol,1996,156:3583
(1999-08-14收稿), 百拇医药
单位:杨俊伟 南京军区南京总医院 解放军肾脏病研究所(南京,210002);龚如军 南京大学医学院博士研究生
关键词:趋化因子;Th1/Th2细胞;免疫应答
T细胞在胸腺中生长发育成初始型T细胞
T细胞在胸腺中生长发育成初始型T细胞(naive T cell)后,通过循环流经全身血液及次级淋巴器官,搜寻抗原呈递细胞(antigen-presenting cell,APC)呈递的抗原。初始型T细胞发现其特异抗原后,即增殖分化为具有新的迁移能力的效应T细胞和记忆T细胞[1]。效应T细胞能够进入外周组织,识别抗原,产生炎性或变态反应;记忆T细胞除具有向外周组织迁移的能力外,还能迁移至淋巴结,进一步刺激产生新一批效应T细胞。因此,T细胞的功能与其迁移能力密切相关,其中必然存在特定的分子机制,使得相应的细胞能够迁移至相应的组织中去。
, 百拇医药
1 趋化因子及其受体是调控白细胞定位的重要成分[2]
白细胞向组织迁移时,在两个阶段受到调控。首先,白细胞在血管内皮表面的滞留及牢固粘附;其次,白细胞从内皮向间质并进一步向特定微环境的迁移。趋化因子在这两个过程中发挥了重要作用。白细胞与内皮的结合牵涉到选择素介导的滚动和整合素介导的牢固粘附。内皮细胞表面的趋化因子与其位于白细胞表面的相应受体相互作用触发了整合素的活化,导致牢固粘附。外周各种组织细胞生成的趋化因子驱使白细胞穿过组织移行至特定微环境。因此,选择素、趋化因子受体和整合素的不同表达为白细胞移行提供了一套“地址码”(addresscode),使白细胞能够在特定位置出现,定位于特定组织。
从上述过程不难得出如下假说:白细胞通过一种多步骤导航模式(multistep navigation mode),对一系列趋化因子先后产生反应。即借助一系列不同的趋化因子受体,使得白细胞能够完成由内皮经间质定位于目的微环境的过程。因此,有些细胞因子在滞留阶段发挥作用,有些则在后继各阶段起作用。显而易见,由于趋化因子及其受体的丰富多样性,对于不同的白细胞,趋化系统能够灵活准确地提供相应的迁移形式。
, 百拇医药
尽管,人们早就认识到选择素和整合素在初始型T细胞和效应/记忆T细胞上表达是不同的,但直到最近,才发现随着抗原刺激程度的不同以及分化程度的不一,许多趋化因子受体在T细胞上的表达也是不一样的。新近的研究表明,趋化因子与受体是调节T细胞及其伴随细胞定位、进一步致敏,以及调节Th1/Th2介导免疫应答所必需的。
2 趋化因子受体在T细胞上的表达
2.1 初始型T细胞和记忆T细胞趋化因子受体表达[3~8] T细胞趋化的早期研究中,人们发现只有记忆/活化T细胞对已知的趋化因子有反应,而初始型T细胞则无反应;而且,淋巴细胞在淋巴组织中的移行可以被百日咳毒素通过抑制趋化因子受体信号传递的作用而阻断。因此,早期研究仅认为只有记忆或活化T细胞才表达Burkitt淋巴瘤受体-1(BLR-1)、C-X-C亚族趋化因子受体5(CXCR5)及CXCR3等趋化因子受体,并对正常T细胞表达和分泌、活化时表达下降的因子(reduced upon activation,normal texpressed and secreted,RANTES)、单核细胞趋化蛋白-1(MCP-1)、-2、-3有反应。
, http://www.100md.com
近年来,人们对趋化因子的功能特性有了进一步认识。许多研究发现趋化因子大体可分为两大类:①炎症性趋化因子,外周组织炎症状态下可被诱导产生或被强烈上调表达;②构成性趋化因子,履行看家功能(housekeeping function),也可能参与构成性的白细胞移行。人类发现的第一个趋化因子即为炎症型,因此只有记忆/活化T细胞表达其同族受体,并对其有反应。最近,人们发现初始型T细胞上也表达趋化因子受体,包括CXCR4、C-C亚型趋化因子受体7(CCR7)。此外,初始型T细胞还表达一种与树突状细胞趋化因子-1(DC-CK1)作用有关的受体,但其结构和功能目前尚不清楚。
T细胞表面趋化因子受体从构成型向炎症型的转变,是T细胞活化、分化的结果,同时它也将改变T细胞的迁移习性。
2.2 Th1、Th2细胞趋化因子受体的表达 与初始型T细胞不同,抗原刺激过的T细胞是不均一的,从另一方面也反应了T细胞活化状态和分化程度的不同。T细胞应答的中心过程是T细胞向Th1型或Th2型分化[9]。T细胞致敏阶段细胞因子的不同组合决定了效应T细胞所产生的细胞因子的类型。CD4+Th1能够产生γ干扰素(IFN-γ),活化单核-巨噬细胞,因此可以杀伤细胞内微生物。相反,CD4+Th2细胞则产生IL-4和IL-5,并与IgE、嗜酸和嗜碱粒细胞主导的反应相关。在MHC-I限制的CD8+T细胞应答中也观察到Th1、Th2的分化。作为Th1、Th2不同效应能力的一部分,它们可能表达不同的趋化因子受体,使得它们向不同组织趋化。
, http://www.100md.com
2.2.1 Th2细胞特异表达的趋化因子受体[10~11] CCR3是嗜酸粒细胞趋化因子(eotaxin)、eotaxin2、RANTES、MCP-2,-3,-4的受体,最初在嗜酸/嗜碱细胞中发现。Th2细胞上选择性地表达CCR3。使用CCR3特异的单克隆抗体检测发现,成人外周血小部分记忆/效应T细胞有CCR3分布,这些细胞同时表现为CD45R0-;分选出这些细胞,体外扩增后发现能产生高浓度的IL-4,IL-5,从而证明这些细胞大多为Th2细胞。体外分化的Th2细胞系也表达CCR3,但Th1则未见表达。此外,不论CD4+还是CD8+T细胞群,CCR3+细胞组分与生成IL-4的细胞数均相近。因此,CCR3+的T细胞可以产生IL-4,而生成IL-4是Th2细胞的重要特征。
目前还不清楚CCR3是否是决定Th2细胞迁移的主要趋化因子受体。除CCR3外,Th2细胞还表达CCR4(TARC的受体),并且在加入胸腺和活化调节的趋化因子(TARC)后,分离出能够动员Ca2+的T细胞,发现这些细胞也能产生IL-4。但CCR4并非Th2的特异标记,它的表达有时可能和产生IL-4的表型相分离;如有文献报道在转化生长因子β(TGF-β)作用下,T细胞系虽然能表达CCR4但并不产生IL-4或IFN-γ。因此只有CCR3才始终与Th2产生IL-4的表型相一致。
, 百拇医药
2.2.2 Th1细胞特异表达的趋化因子受体[11~14] 与Th2细胞能够选择性地表达趋化因子受体一样,Th1也选择性地表达某些受体。CXCR3就是其中之一,它是趋化因子IP-10(IFN-γ诱导蛋白-10)、Mig(IFN-γ诱导的单核因子)和I-TAC(IFN-γ可诱导的T细胞α趋化诱导剂)的受体,所有这些趋化因子均可被IFN-γ上调。CXCR3在大多数外周血记忆T细胞都有表达,但Th1表达水平远高于Th2,与此相应,Th1对IP-10反应所需的剂量仅为Th2的十分之一。
CCR5是RANTES、巨噬细胞炎性蛋白-1α(MIP-1α)和MIP-1β的受体,但是否为Th1的特异性受体尚有争议。虽然有研究发现CCR5选择性地表达于Th1,但Sallusto等的工作证实,尽管CCR5在Th1上有表达,而Th2没有表达,但在新近活化的T细胞,不管其功能、分化如何都有同等量的CCR5表达。使用CCR5特异性抗体,发现CCR5存在于一小部分CD26+、L-选择素(L-selectin)、CD45R0+的外周血T细胞,而这一系列表型均与T细胞的新近活化相一致。此外,当撤减IL-2时,CCR5则迅速消失,而CCR3、CXCR3却无变化。因此CCR5+T细胞可能仅代表最新产生的循环效应细胞。
, http://www.100md.com
尽管现有资料十分有限,但已有的研究发现Th1、Th2表达不同的趋化因子受体,因而对趋化因子反应也不相同。近期对小鼠淋巴系统的研究也已观察到类似现象。
3 IFN-α和TGF- β调节趋化因子受体的表达[11,15~17]
T细胞的分化不仅受抗原刺激强度的调节,同时还受致敏阶段周围细胞因子的影响。这些促进T细胞分化的细胞因子来源于APC、应答的T细胞或周围旁观细胞。尽管IL-2和IL-4在Th1、Th2分化过程中起着主要作用,但其它细胞因子同样也影响T细胞的最终分化。即便在没有IL-12或仅存在IL-4的环境下,IFN-α仍可诱导人Th1分化。而TGF-β则能够抑制人T细胞分化,使增殖的T细胞处于半初始化(semi-naive)状态。半初始化T细胞除IL-2外,不生成其它任何细胞因子,但在适当刺激后,仍可进一步分化为Th1或Th2。
Sallusto等研究发现,IFN-α和TGF-β除影响分化外,还能显著地影响不同类型趋化因子受体的表达。在Th1或Th2分化环境中加入IFN-α后,CCR3、CCR4的表达受到抑制,而CCR1的表达上调。因此,与IL-12诱导分化的Th1相比,IFN-α诱导分化的Th1细胞还上调表达有大量的CCR1,而CCR1能使T细胞对更多的趋化因子作出反应。相反,TGF-β抑制CCR2、CCR3和CCR5的表达,而上调CCR4、CCR7的表达。所以,与其抑制分化的作用相一致,那些能促进T细胞向外周组织迁移的趋化因子受体也受到TGF-β的抑制;而有助于T细胞归巢的趋化因子受体则得到增强表达。总之,IFN-α、TGF-β能够调节T细胞趋化因子受体的表达,灵活地调节着白细胞的运动,对特定组织中细胞的组成也有深刻的影响(见图1)。
, http://www.100md.com
图1 T细胞趋化因子受体表达的调节
DC:树突状细胞 DC-DK1-R:树突状细胞趋化因子1受体
Naive T cell:初始型T细胞 DTH:迟发型超敏反应
4 趋化因子协调次级淋巴器官中T细胞、B细胞和树突状细胞的关系[18]
趋化因子及其受体在引导特定细胞迁移至特定炎症组织及淋巴器官,诱发初级免疫应答产生的过程中也发挥了重要作用(见图2)。
淋巴结中初始型T细胞和树突状细胞(DC)充分相互作用,是T细胞致敏的重要场所。未成熟的DC及单核细胞(DC的直接前体),表达多种受体,如CCR1、2、5、6,迅速募集到外周炎症组织。进入炎症组织后,DC立即俘获抗原,同时受炎症因子,如TNF-α、IL-1或细菌病毒产物(如LPS或dsRNA等)的刺激而分化成熟。DC的成熟不仅出现粘附分子或协同刺激分子上调,而且可使之向次级淋巴器官迁移。LPS或TNF-刺激DC产生大量炎症型趋化因子,进一步募集未成熟的DC;并可使其自身的趋化因子受体下调,有利于DC及时从炎症组织中退出。当DC的炎症型趋化因子受体下调时,构成性趋化因子受体如CCR4、CCR7、CXCR4在成熟的DC得到上调表达。CCR7对引导成熟的DC进入淋巴管尤为重要,这是因为淋巴管内皮细胞能够产生CCR7的同族配体——次级淋巴组织趋化因子(SLC)。DC从淋巴管引流到局部淋巴结,淋巴结中固有的成熟DC可产生另一种CCR7配体EBI1配体趋化因子(ELC),控制DC定位于淋巴结的T细胞区。
, http://www.100md.com
图2 趋化因子及其受体在T细胞致敏和Th1/Th2介导的免疫应答中的作用
APC:抗原呈递细胞 naive T:初始型T细胞 SLC:次级淋巴样组织趋化因子
DC:树突状细胞 Mig:γ-干扰素诱导的单核因子 ELC:EBI1配体趋化因子
MIP:巨噬细胞炎性蛋白DCA-1:B细胞趋化因子1MDC:巨噬细胞来源的趋化因子
SDF-1:基质细胞来源的因子1PSGL-1:P-selectin糖蛋白配体1IP-10:干扰素可诱导蛋白10
与DC相伴的初始型T细胞,通过另一条途径——高内皮细胞的毛细血管后微静脉(HEV)进入淋巴结。L-selectin和淋巴细胞功能相关抗原-1(LFA-1)在该过程中起促进作用。淋巴组织中至少产生包括基质细胞来源的因子-1(SDF-1)、SLC、ELC在内的三种趋化因子,可以使血循环中T细胞迅速滞留于细胞间粘附分子-1(ICAM-1)表达区域,而它们相应的受体CXCR4和CCR7在初始型T细胞均呈高水平表达。因此,CXCR4和CCR7同时也可能是介导牢固粘附和穿过HEV迁移的重要趋化因子受体。初始型T细胞进入淋巴结后,自由地渗滤通过没有细胞外基质的副皮质区,使之与DC充分作用。DC自身也分泌ELC或DC-CK1等趋化因子,除了对初始型T细胞有活性,吸引T细胞外,还可能在T细胞致敏的起始步骤中起作用。
, 百拇医药
初始型B细胞,与初始型T细胞一样,也表达CXCR4和CCR7,从HEV进入淋巴结后定位于滤泡B细胞区。初始型B细胞还表达CXCR5,通过基因打靶技术业已证实CXCR5是淋巴滤泡及生发中心形成所必需的成份。人类CXCR5的配体是B细胞趋化因子-1(BCA-1),小鼠中的相应物质为B淋巴细胞趋化剂(BLC),它们均由滤泡中的细胞产生,在吸引初始型B细胞以及活化的Th细胞、形成滤泡结构的过程中发挥作用。活化的B细胞也能生成巨噬细胞衍生的趋化因子(MDC)等趋化因子,可能促进T细胞和B细胞的相互作用。
5 趋化因子影响白细胞迁移的作用
5.1 迟发型超敏反应和变态反应中白细胞迁移的调节[11,13,19] 局部组织的炎症增强了白细胞内皮的粘附和移行。这一现象的基础是内皮细胞粘附分子的上调以及内皮细胞和组织细胞分泌多种炎症型趋化因子,这些物质分布的差异有可能决定哪些细胞将会优先聚集到某一特定的炎症部位(见图2)。
, http://www.100md.com
在迟发型超敏反应的组织中,炎症性细胞因子,如TNF-α、IL-1和IFN-γ,能够促进内皮细胞表达E-selectin、P-selectin和血管细胞粘附分子-1(VCAM-1),并刺激产生多种趋化因子,包括IP-10、Mig。Th1表达P-selectin糖蛋白配体-1(PSGL-1),可与E-selectin、P-selectin结合,涉及T细胞移行到迟发型超敏反应区。Th2上虽然也曾发现PSGL-1,但它并不与E-selectin、P-selectin结合,有研究显示,它可能与变态反应组织内皮上优先表达的某种未知配体功能相关。
最近的研究表明,有些趋化因子受体并不依赖于整合素的作用,是直接在粘附过程中发挥作用的。NK和一些活化T细胞上均表达CX3CR1,因此可与fractalkine(内皮细胞上一种整合性膜趋化因子)相作用而介导滚动、粘附。CCR1也有类似作用,它可将E-selectin、P-selectin作为替代配体与之结合。此外有研究发现,除LFA-1外,记忆/活化T细胞还表达很晚出现的抗原-4(VLA-4)和其它整合素;虽然VLA-4与其配体VCAM-1的结合不需要趋化因子的参与,但是,VLA-4与纤维连接蛋白的作用却必须要有趋化因子参与,而且纤维连接蛋白在T细胞移行到组织中起了重要作用。尽管人们目前尚不清楚IP-10和Mig,哪一个在粘附或移行水平发挥作用,但由于它们的表达是受调节的,且其同族受体CXCR3在Th1上呈高水平的表达,因此这些趋化因子可能在选择性地募集Th1至以IFN-γ为主的炎症部位起重要作用。此外,由于T细胞均表达高水平的炎症性趋化因子相应的同族受体,如CCR5,因此RANTES、MIP-1α和MIP-1β还可以吸引新近活化的T细胞。
, 百拇医药
与变态反应关系最为密切的趋化因子为嗜酸性粒细胞趋化因子(eotaxin),它在肺中生成,并在肺中保持高浓度。eotaxin仅与CCR3结合,而Th2上选择性地表达CCR3。因此,eotaxin可以募集Th2及其它炎症细胞到变态反应组织中。除eotaxin外,其它趋化因子也可能参与了变态反应,在变态反应不同阶段,募集不同的效应细胞。
5.2 趋化因子受体在白细胞共定位中的作用[14] 我们常可以通过一个T细胞的伴随细胞来判断该T细胞的种类。Th2常伴随有嗜酸和嗜碱性细胞,而Th1则更常与巨噬细胞和中性粒细胞共处。这可能是由于抗原特异性T细胞及其它不同类型的效应细胞共享相同的趋化因子受体,而使它们共处一处。例如,Th2、嗜酸、嗜碱细胞均普遍表达CCR3。这三种细胞能够协同作用产生变态炎症反应,但更重要的是嗜酸、嗜碱细胞的活化、存活依赖于Th2分泌的IL-5,IL-4。再如,Th1细胞和单核细胞均普遍表达CCR1和CCR5,单核细胞是巨噬细胞和DC的前体。这就使得APC、清道夫细胞和炎症T细胞能同时募集到迟发型超敏反应组织中。
, 百拇医药
对炎症组织中T细胞的趋化因子受体的直接分析已证实了上述理论。因此,在变态反应,如鼻息肉或接触性皮炎都有CCR3+的T细胞、大量嗜酸/嗜碱性粒细胞浸润;而迟发型超敏反应,如类风湿关节炎,则仅有CCR5+和CXCR3+的T细胞浸润。类风湿关节炎时,滑膜液中有大量的CCR5+T细胞,而血液及淋巴组织中的数量则相对很少,这从另一方面说明T细胞定位是有选择的。
NK细胞是先天免疫的重要效应细胞,目前对NK在体内移行的途径知之甚少。NK细胞表达多种趋化因子受体,能使其和其它白细胞一起成为机体的首道防线。已有研究报道,NK可对多种趋化因子均有反应,包括IP-10,LT(lymphotactin)、RANTES、MIP-1、MCP-1,2,3、MDC。
结语[20] 大量趋化因子及其受体的发现,使我们进一步认识到淋巴细胞是如何选择性地移行至组织的。目前,我们面临的首要任务是澄清所有趋化因子受体的功能和表达形式。各种受体在不同细胞中的信号传递能力也是一个重要的研究领域,但目前仍知之甚微。一些受体可能采用不同的信号途径,因此,不仅在移行,在淋巴细胞的活化、转化甚至存活中都可能发挥作用。此外,趋化因子受体信号传递能力不同,也可能是由于G蛋白信号系统的调节蛋白表达不同所致;这些蛋白的表达对决定趋化因子受体的功能至关重要。另外一个重要的研究领域是组织中趋化因子生成的调控,现已清楚,免疫反应的性质是由组织中生成的趋化因子的种类所决定的。
, http://www.100md.com
趋化因子及其受体可能是当今治疗炎症疾病最有希望的研究领域。多步骤导航模式,即一个迁移细胞在其移行的不同特定阶段可能表达不同的趋化因子受体、对不同的趋化因子有反应;因此,如果阻断趋化因子受体则会导致迁移细胞迷失方向,不能到达其最终目标。该理论同时也解释了为何阻断人类趋化因子可以非常有效地抑制多种炎症反应。趋化因子的功能是使白细胞选择性地向特定组织移行,所以拮抗趋化因子受体的治疗将比现有疗法更具选择性,产生更少的免疫抑制副作用。目前,已生产的趋化因子受体拮抗剂多种多样,包括单克隆抗体,趋化因子修饰物以及最近研制的小分子无机物。在未来的几年中,它们很可能成为治疗自身免疫疾病和变态反应疾病的有效药物。
总之,趋化因子已从未知的细胞因子的分枝学科发展成为一个涉及生物学许多方面的新研究领域。趋化因子及其受体在生长发育、上皮和内皮细胞移行、神经生物学、T细胞活化和感染性疾病中的作用正逐步为人类所认识。
参考文献
, 百拇医药
1 Mackay CR.Immunological memory.A dv Immunol,1993,53:217
2 Picker LJ,Butcher EC.Physiological and molecular mechanisms of lymphocyte homing.Annu Rev Immunol,1992,10:561
3 Bargatze RF,Butcher EC.Rapid G protein -regulated activation event involved in lymphocyte binding to high endothelial v enules.J Exp Med,1993,178:367
4 Foster R,Emrich T,Kremmer E et al.Ex pression of the G-protein-coupled recept or BLR1 defines mature,recirculating B cells and a subset of T-helper memory cel ls.Blood,1994,84:830
, 百拇医药
5 Loetscher P,Seitz M,Baggiolini M et al.Interleukin-2 regulates CC chemokine receptor expression and chemotactic resp onsiveness in T lymphocytes [see comment s].J Exp Med,1996,184:569
6 Schall TJ,Bacon K,Toy KJ et al.Selec tive attraction of monocytes and T lymph ocytes of the memory phenotype by cytoki ne RANTES.Nature,1990,347:669
7 Yoshida R.Imai Y,Hieshima K et al.Mo lecular cloning of a novel human CC chem okine EBI1-ligand chemokine that is a specific functional ligand for EBI1,CCR7.J Biol Chem,1997,272:13803
, 百拇医药
8 Hieshima K,Imai T,Baba M et al.A novel human CC chemokine PARC that is most homologous to macrophage-inflammatory protein-1 alpha/LD78 alpha and chemotactic for T lymphocytes,but not for monocytes .J Immunol,1997,159:1140
9 杨俊伟,黎磊石.Th1/Th2细胞与免疫.肾脏病与透析肾移植杂志,1995,4:533
10 Ponath PD,Qin S,Post TW et al. Molecular cloning and characterization of a human eotaxin receptor expressed sel ectively on eosinophils [see comments].J Exp Med,1996,183:2437
, http://www.100md.com
11 Sallusto F,Lenig D,Mackay CR et al.Flexible programs of chemokine rece ptor expression on human polarized T hep er 1 and 2 lymphocytes.J Exp Med,1998,18 7:875
12 Luster AD,Ravetch JV.Biochemical characterization of a gamma interferon- inducible cytokine(IP-10).J Exp Med,1987 ,166:1084
13 Bonecchi R,Bianchi G,Bordignon P P et al.Differential expression of che mokine receptors and chemotactic respons iveness of type 1 T helper cells(Thls)and Th2s.J Exp Med,1998,187:129
, 百拇医药
14 Loetscher P,Uguccioni M,Bordoli L et al.CCR5 is characteristic of Th1 lymphocytes [letter].Nature,1998,391:344
15 刘俊铎,刘志红.Th1/Th2细胞与自身免疫性疾病.肾脏病与透析肾移植杂志,1997,6:555
16 Cella M,Scheidegger D,Palmer Leh mann K et al.Ligation of CD40 on dendr itic cells triggers production of high l e vels of interleukin-12 and endhances T cell stimulatory capacity:T-T help via A PC activation.J Exp Med,1996,184:747
, 百拇医药
17 Rogge L,Barberis-Maino L,Biffi M et al.Selective expression of an inte rleukin-12 receptor component by human T helper 1 cells.J Exp Med,1997,185:825
18 Sozzani S,Sallusto F,Luini W et al.Migration of dendritic cells in res ponse to formyl peptides,C5a,and a disti nct set of chemokines.J Immunol,1995,155 :3292
19 Taub DD,Lloyd AR,Conlon K et al .Recombinant human interferon-inducible protein 10 is a chemoattractant for hum an monocytes and T lymphocytes and promo tes T cell adhesion to endothelial cells .J Exp Med,1993,177:1809
20 Strieter RM,Standiford TJ,Huffin agle GB et al.“The good,the bad,and the ugly.”The role of chemokines in models of human disease.J Immunol,1996,156:3583
(1999-08-14收稿), 百拇医药