�����ݸ˾�cagA�����������ѡ��ԭ��Tϸ��Fas Ligand�鵼�����ĵ�������
���ߣ����̵� ���� ��ʤ �ܵ�Ԫ �ֻ���
��λ�����ݣ���һ��ҽ��ѧ�Ϸ�ҽԺȫ���������о���510515
�ؼ��ʣ��ݸ˾������ţ����ߣ�ϸ������������
�л�ҽѧ��־001105
��ժҪ�� Ŀ�� ���������ݸ˾�(Hp)���ֹ��ܵ�����Tϸ����������������̽��������Fas/FasL�鵼�Ļ���Tϸ�������е����ã�ΪHp��������Ѱ��ȫ��Ч������ԭ������ Ӧ��JAM������cagA����Hp��ͬԴ����ͻ���ꡢ��������ø��ճ����HpaA����Ĥ����Hop25��35��38��Tϸ��ϵ���������������˱Ƚϣ�Ӧ����ʽϸ���������������ر���cagA���Ծ����յ�Tϸ��FasL��������������˼�⣻Ӧ�ý�������ø���Ƽ��͵��ϳ����Ƽ��ֱ���ߺͽ���Hp�鵼��Tϸ����������֤ʵ����FasL�ϳɵĹ�ϵ����� Hp cagA���������������Tϸ������FasL�������յ�Tϸ��������������þ�ǿ��ͬԴ����ͻ���ꡣHp��ԭ��Ʒ������ø��Tϸ����ɱ����������Hop38������ǿ��Hop25��Hop35��HpaA�����ý�������������ø���Ƽ������cagA����Hp��Tϸ�������������������ϳ����Ƽ�Emetine��ͨ������Tϸ��FasL�����������Hp�鵼��Tϸ������������ cagA����Hop38��ͨ������FasL�������Tϸ�����ܣ�����ø����Ĥ����Hop25��Hop35��HpaA����ͨ������Tϸ�����������ܻ��Ƶķ���������ɰ�ȫӦ�����������ơ�
, http://www.100md.com
The evaluation of cagA and other vaccine candidate proteins of Helicobacter pylori on T cell apoptosis mediated by Fas/Fas ligand interaction
WANG Jide CHEN Ye LAI Zhuosheng
(PLA Institute for Digestive Disease, Nanfang Hospital, First Military Medical University, Guangzhou 510515, China)
��Abstract�� Objective To evaluate the effect of several surface proteins of Helicobacter pylori (H. pylori) on inducing apoptosis of T cells through the modulation of Fas/Fas ligand (FasL) interaction and to provide information on the screen of safe and effective immunogen for H. pylori vaccine. Methods T cell apoptosis induced by cagA+ and isogenic cagA knockout strain, recombinant urease, HpaA and outer membrane proteins, Hop25, Hop35 and Hop38 was detected by JAM assay. Flow cytometry was used to evaluate the increase in FasL expression on T cells under the stimulation of these bacterial proteins. Metalloprotease inhibitor, 1, 10-Phenanthroline (PHE) and protein synthesis inhibitor, emetine, were used to change the activity of FasL to confirm the association of FasL with H. pylori mediated killing toward T cells. Results The effect of cagA+ H.pylori was stronger in both the induction of apoptosis and FasL expression of T cells than that of cagA knockout strain. H. pylori urease could not kill T cell, while Hop38 had the strongest effect among the tested H. pylori proteins on the induction of T cell apoptosis through increasing FasL expression. Metaloprotease inhibitor could increase the killing of T cells induced by H. pylori because of its ability to prevent the cleavage of membrane-bound FasL protein. On the other hand, emetine could decrease H. pylori mediated killing by inhibiting of FasL synthesis on T cells. Conclusion cagA and Hop38 protein might be involved in the negative regulation of T cell growth through upregulating FasL expression urease and other Hop proteins tested might be safe and effective to be used as the antigens for H. pylori vaccine.
, ��Ĵҽҩ
��Key words�� Helicobacter pylori�� Immunity, cellular�� Apoptosis
�����ݸ˾�(Helicobacter pylori, Hp)��Ⱦ��������θ�������������ܴٽ�θ���ķ��������õ�����ҩ�����Ʒ������ڲ��������Բ�������ô��ϸ���������ҩ�Ȳ��㣬��ˣ���չ���������ʮ����Ҫ��
Hp��Ⱦ�ɵ�����������Tϸ�����ܡ���������θճĤ������Hp����Tϸ����¡[1]�����ھֲ�������Ѫ�м���˿�Hp���壬���������²����������ܻ��ƽ������о�[2��3]����Ϊ����ø������Ҫ�Ķ�ֲ���²�����֮һ��cagA�²�������(Pathogenicity Island, PAI)�����һϵ����ϸ������ת���йصĶ���[3]����Hp�����²����������ܵĻ����в����ˣ�����֤�����Ч���Ͱ�ȫ�ԡ�����Hp�ܴ̼�Tϸ������Fas Ligand (FasL)��ͨ��Fas/FasL����ö���ɱ����ɱ�ˣ���Ҳ����Hp������Tϸ����Ӧ�Խ���Bϸ�����ܵĿ��ܻ��ơ��ڱ��о��У�����Ӧ������Tϸ��ϵģ��������Hp���������ѡ��ԭ��Fas/FasL�鵼Tϸ�������ĵ������ã���Ԥ�������������߱���ʱ�Ŀ����ԡ�
, ��Ĵҽҩ
�����뷽��
һ��Hp�����Tϸ��ϵ
Hp���ʱ���ATCC26695����cagA�²���ͬԴͻ����D-1�������¿���˹��ѧErnst���ڻ���������37�棬��������������������ΪSkirrowѡ������������Һ��Ϊ����������CD4����Tϸ��ϵCem-C7 (Th2����)��Jurkat�������������ռ�������������ģ�����RPMI1640Һ������
����ϸ����������
�����ױ���[4]Ӧ��JAM�����ⶨTϸ��������3H��ǵ�Tϸ����Hp�����������干ͬ����24 h(ϸ��/ϸ������Ϊ1 000��1������1��4ϡ��)���ѽⲢ�ռ�ϸ��DNA����Һ���ⶨ�����ݶ�ʧDNA�ı���������Hp��ɱ���ʡ��ڲⶨHp��ԭ��ɱ������ʱ��Hp��ԭӦ��Ũ�Ⱦ�Ϊ10 ��g/ml��
, ��Ĵҽҩ
����Hp��ԭ
Hp��Ĥ����Hop25��Hop35��Hop38ϵ�ɹ����������ṩ����������ø��ճ����HpaAϵ���дӺ���Hp����ø��������pHP902�ͺ�HpaA��������pHP901�Ĺ��̾��д�����
�ġ���ʽϸ���Ǽ��FasL����
Tϸ����Hp(1��300)���俹ԭ(10 ��g/ml)����24 h���ռ���Ưϴϸ����Ӧ���ÿ���FasL����C-20(Santa Cruz��˾)������FITC�������IgGȾɫ�����տ���Ӧ��������IgG����ʽϸ���Ƿ������Ƚ�Hp��������Ǵ����飬����FasL���������
�塢��������ø���Ƽ���Hpɱ��Tϸ��������
����ϸ�������FasL���ڱ���Һ(��������Һ�е�Ѫ��ɷ�)�еĽ�������ø���Ƽ����⣬��Ϊ������FasL���������Է�����Fas������ϸ����������������Ĥ���FasL������Ӧ�ý�������ø���Ƽ�1,10-phenanthroline(PHE������Sigma��˾)���Fas�鵼��Tϸ����������[5]����ǰ��JAM�����У����Tϸ����Hp����ʱ����Һ����2.5 mmol/L��PHE������ɱ���ʵĸı䡣
, ��Ĵҽҩ
�������ϳ����Ƽ���Hpɱ��Tϸ����FasL�������������
Ϊ̽��Hpɱ��Tϸ���Ƿ���Ҫ�ºϳɵ�FasL���ף�����Ӧ�õ��ϳ����Ƽ�emetine( ����Sigma��˾)����Tϸ��FasL�ϳɼ�Hp�鵼���µ�������[6]��10 mmol/L emetine��Tϸ������45 min����Hp����FasL�����Ӱ�켰�յ�������������
�ߡ�ͳ��ѧ����
���ֵ���(ɱ��)�ʾ���Sigma plot 3.0��ͼ����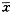 ��s��ʾ��������student's t���顣P<0.05Ϊ�������������塣
��s��ʾ��������student's t���顣P<0.05Ϊ�������������塣
���
һ��cagA��Hp��Tϸ�������Ĺ�ϵ
, ��Ĵҽҩ
cagA����Hp�������ǻ�������������������ɱ��Tϸ��Cem-C7��ɱ������43%��53%֮�䣬��CagͬԴͻ����D-1��ɱ����С��10%(P<0.05)����Jurkat��ɱ����ͬ����𣬵�ɱ���ʵ���30%(��1)��
��1 cagA���Լ����Ծ�����Tϸ�������ʱȽ�(%����s) Tϸ��
ATCC 26695
���
D-1���
ATCC 26695
����
D-1����
, http://www.100md.com
Cem-C7
43��4��
9��4
53��5��
4��2
Jurkat
26��2
6��2
28��4
6��3
����D-1��ȣ�P<0.05
����Hp��ԭ�����Tϸ�������������Ƚ�
, http://www.100md.com
��cagA����Hp�����Ϊ���գ����DZȽ��˼���Hp���浰����Tϸ��Cem-C7���������á��������cagA������ATCC26695�����ճ����HpaA����Ĥ����Hop25��Hop35��Hop38��ɱ���ʷֱ�Ϊ49.0��0.4��18��2��9��4��22��5��75��3��������ø�����á�����Hop38�Ķ�����ǿ��ATCC26695����� Hop25��HpaA�����ý�����Ӧ��Jurkatϸ����ͬ�����֡�
����Hp��������ֶ�Tϸ��FasL����ĵ�������
��ʽϸ�������� Hop38��cagA�����Դ̼�Tϸ������FasL��ͼ1ʾcagA����Hp��ATCC26695�����Tϸ������FasL����cagA����ͻ����D-1�������á��˽����JAM���һ�¡�
�ġ���������ø���Ƽ�������
Ӧ��JAM���������Ƿ����ڽ�������ø���Ƽ�PHE���ڵ�����£�������ATCC26695������������������Cem-C7��Jurkat��ɱ�����þ������(P<0.05)��˵����������ø���Ƽ�������Ĥ���FasL��ɱ�������������ͼ2��
, ��Ĵҽҩ
ͼ1 cagA����Hp���Tϸ��FasL�����Ӱ��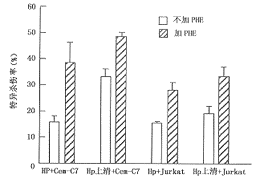
ͼ2 ��������ø���Ƽ���Hp�յ���Tϸ��������Ӱ��
�塢���ϳ����Ƽ�����Hp�鵼��FasL���PTϸ������
10 mmol/L emetine�ܲ����������ϸ���ϳ��µ��ף���ͼ3��ʾ���������µ�Hp26695�鵼��Tϸ��FasL����,ͬʱ����Hp��Cem-C7��ɱ������(ͼ4)��˵��Hp��Tϸ��������Ҫ�ºϳɵ�Fas L���ף�������ֱ��������Tϸ�������Fas���塣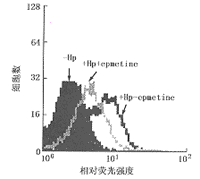
ͼ3 ���ϳ����Ƽ�����Hp�յ���
, http://www.100md.com Tϸ��FasL����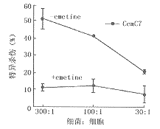
ͼ4 ���ϳ����Ƽ�����Hp��
cem-c7��ɱ������
����
�ڻ����������ԭ�����߷�Ӧ�У�Tϸ����ֱ�Ӳ���ϸ�����߲�����Bϸ���������壬��������ԭ����������Ҳ����Tϸ���鵼�������о����֣�Hp��ȾʱθTϸ����ֳ����Ծ����Hp���������Tϸ����ֳ(����Hp��ԭ���⣬������ø�ɴ̼�Tϸ����ֳ)[7]�����ǻ���Hp��������Tϸ���������书�ܣ��Ӷ�ʹ��Ⱦ����������
�˽�Hp���ܺͱ��濹ԭ��Tϸ��������Ӱ���ܹ�Ԥ������Ϊ����ԭʱ��Ч���Ͱ�ȫ�ԡ�����ø����Tϸ�����������ٽ�����ֳ���Ӷ���������ȫ����Ч������ԭ��cagA�� Hop38��Tϸ�������������ʵ��Ӧ���п������ܴ̼�Tϸ�������㹻��Ч�ĸ���ϸ�����Ӷ����ܻ�ñ�������������������塣HpaA�� Hop 25������������Tϸ�����������ã�������������������á�
, ��Ĵҽҩ
��Ȼ�б���Ӧ��cagA��������С����Ա������ﲻ��cagA����Hp����ĸ�Ⱦ[8]�������о�������һЩ���ֲ���֧��cagA�������ߵ���Чǰ����cagA���Ծ������������֢��/�����صĻ��ߣ���Щ����θ����Ƥ��AP-1��NFkB��ת¼���ӻ�������[9]���伤��ɵ��¶�����֢����������ئ�(IFN-��)������2(IL-2)�������������Ӧ�(TNF-��)��IL-8�ȵı���Ӷ��Ӿ�θճĤ��[10]�����������Tϸ��Fas L���������������������2��NFkB�Ľ��λ�㣬3��AP-1���λ��[11]�����cagA����ͨ������ת¼���Ӷ�����Tϸ��FasL�������������Fas���Ե�θ����Ƥ�����ܴٽ�Tϸ��֮�以���ɱ���Ӿ�����Hp��ԭ����Ӧ�Ժ��������ܡ����о�����cagA���������Ծ����ڼ���FasL��ɱ��Tϸ������ľ����Ϊ������һ�����ṩ���µ�֤�ݡ�
����Fas/ FasL��Hp��ԭ���µ�Tϸ�������е����ã�����T����ϸ���������ֹ��ܱ��ͣ�Th1������IFN-�á�TNF-���ȹ㷺������Hp��Ȼ��Ⱦ��θճĤ�������߱�����Ч�Ķ�����θճĤ��Th2ϸ��Ϊ�����Ҿݱ����������ϸ����Fas/ FasL�鵼ϸ�������������Բ�ͬ����һ�����о�Ӧ�������������Щ�����ѡ��ԭ�Բ�ͬTϸ�����͵����ã��������տ˷�Tϸ�����ܣ���ð�ȫ��Ч��Hp������ԭ��
, ��Ĵҽҩ
������Ŀ��������Ȼ��ѧ����������Ŀ(39700007)
�����
1��D'Elios MM, Manghetti M, Almerigogna F, et al. Different cytokine profile and antigen-specificity repertoire in Helicobacter pylori-specific T cell clones from the antrum of chronic gastritis patients with or without peptic ulcer. Eur J Immunol, 1997,27:1751-1755.
2��Doig P, Trust TJ. Identification of surface-exposed outer membrane antigens of Helicobacter pylori. Infect Immun, 1994,62:4526-4533.
, ��Ĵҽҩ
3��Akopyants NS, Clifton SW, Kersulyte D, et al. Analyses of the cag pathogenicity island of Helicobacter pylori. Mol Microbiol, 1998,28:37-53.
4��Matzinger P. The JAM test. A simple assay for DNA fragmentation and cell death. J Immunol Methods, 1991,145:185.
5��Mitsiades N, Poulaki V, Kotoula V, et al. Fas ligand is present in tumors of the Ewing's sarcoma family and is cleaved into a soluble form by a metalloproteinase. Am J Pathol, 1998,153:1947-1956.
, ��Ĵҽҩ
6��Li JH, Rosen D, Ronen D, et al. The regulation of CD95 ligand expression and function in CTL. J Immunol, 1998,161:3943-3949.
7��Haeberle HA, Kubin M, Bamford KB, et al. Differential stimulation of interleukin-12 (IL-12) and IL-10 by live and killed Helicobacter pylori in vitro and association of IL-12 production with gamma interferon-producing T cells in the human gastric mucosa. Infect Immun, 1997,65:4229-4235.
8��Marchetti M, Rossi M, Giannelli V, et al. Protection against Helicobacter pylori infection in mice by intragastric vaccination with H. pylori antigens is achieved using a non-toxic mutant of E. coli heat-labile enterotoxin (LT) as adjuvant. Vaccine, 1998,16:33-37.
, ��Ĵҽҩ
9��Glocker E, Lange C, Covacci A, et al. Proteins encoded by the cag pathogenicity island of Helicobacter pylori are required for NF-kappa B activation. Infect Immun, 1998,66: 2346-2348.
10��Yasumoto K, Okamoto S, Mukaida N, et al. Tumor necrosis factor and interferon synergistically induce interleukin 8 production in a human gastric cancer cell line through acting concurrently on AP-1 and NF-kappaB-like binding sites of the interleukin 8 gene. J Biol Chem, 1992,267: 22506-22511.
11��Matsui K, Fine A, Ahu B, et al. Identificaiton of two NF-kappa B sites in mouse CD95 ligand (Fas ligand) promoter: Functional analysis in T cell hybridomas. J Immunol, 1998,161: 3469-3473.
(�ո����ڣ�1999-11-25), ��Ĵҽҩ
��λ�����ݣ���һ��ҽ��ѧ�Ϸ�ҽԺȫ���������о���510515
�ؼ��ʣ��ݸ˾������ţ����ߣ�ϸ������������
�л�ҽѧ��־001105
��ժҪ�� Ŀ�� ���������ݸ˾�(Hp)���ֹ��ܵ�����Tϸ����������������̽��������Fas/FasL�鵼�Ļ���Tϸ�������е����ã�ΪHp��������Ѱ��ȫ��Ч������ԭ������ Ӧ��JAM������cagA����Hp��ͬԴ����ͻ���ꡢ��������ø��ճ����HpaA����Ĥ����Hop25��35��38��Tϸ��ϵ���������������˱Ƚϣ�Ӧ����ʽϸ���������������ر���cagA���Ծ����յ�Tϸ��FasL��������������˼�⣻Ӧ�ý�������ø���Ƽ��͵��ϳ����Ƽ��ֱ���ߺͽ���Hp�鵼��Tϸ����������֤ʵ����FasL�ϳɵĹ�ϵ����� Hp cagA���������������Tϸ������FasL�������յ�Tϸ��������������þ�ǿ��ͬԴ����ͻ���ꡣHp��ԭ��Ʒ������ø��Tϸ����ɱ����������Hop38������ǿ��Hop25��Hop35��HpaA�����ý�������������ø���Ƽ������cagA����Hp��Tϸ�������������������ϳ����Ƽ�Emetine��ͨ������Tϸ��FasL�����������Hp�鵼��Tϸ������������ cagA����Hop38��ͨ������FasL�������Tϸ�����ܣ�����ø����Ĥ����Hop25��Hop35��HpaA����ͨ������Tϸ�����������ܻ��Ƶķ���������ɰ�ȫӦ�����������ơ�
, http://www.100md.com
The evaluation of cagA and other vaccine candidate proteins of Helicobacter pylori on T cell apoptosis mediated by Fas/Fas ligand interaction
WANG Jide CHEN Ye LAI Zhuosheng
(PLA Institute for Digestive Disease, Nanfang Hospital, First Military Medical University, Guangzhou 510515, China)
��Abstract�� Objective To evaluate the effect of several surface proteins of Helicobacter pylori (H. pylori) on inducing apoptosis of T cells through the modulation of Fas/Fas ligand (FasL) interaction and to provide information on the screen of safe and effective immunogen for H. pylori vaccine. Methods T cell apoptosis induced by cagA+ and isogenic cagA knockout strain, recombinant urease, HpaA and outer membrane proteins, Hop25, Hop35 and Hop38 was detected by JAM assay. Flow cytometry was used to evaluate the increase in FasL expression on T cells under the stimulation of these bacterial proteins. Metalloprotease inhibitor, 1, 10-Phenanthroline (PHE) and protein synthesis inhibitor, emetine, were used to change the activity of FasL to confirm the association of FasL with H. pylori mediated killing toward T cells. Results The effect of cagA+ H.pylori was stronger in both the induction of apoptosis and FasL expression of T cells than that of cagA knockout strain. H. pylori urease could not kill T cell, while Hop38 had the strongest effect among the tested H. pylori proteins on the induction of T cell apoptosis through increasing FasL expression. Metaloprotease inhibitor could increase the killing of T cells induced by H. pylori because of its ability to prevent the cleavage of membrane-bound FasL protein. On the other hand, emetine could decrease H. pylori mediated killing by inhibiting of FasL synthesis on T cells. Conclusion cagA and Hop38 protein might be involved in the negative regulation of T cell growth through upregulating FasL expression urease and other Hop proteins tested might be safe and effective to be used as the antigens for H. pylori vaccine.
, ��Ĵҽҩ
��Key words�� Helicobacter pylori�� Immunity, cellular�� Apoptosis
�����ݸ˾�(Helicobacter pylori, Hp)��Ⱦ��������θ�������������ܴٽ�θ���ķ��������õ�����ҩ�����Ʒ������ڲ��������Բ�������ô��ϸ���������ҩ�Ȳ��㣬��ˣ���չ���������ʮ����Ҫ��
Hp��Ⱦ�ɵ�����������Tϸ�����ܡ���������θճĤ������Hp����Tϸ����¡[1]�����ھֲ�������Ѫ�м���˿�Hp���壬���������²����������ܻ��ƽ������о�[2��3]����Ϊ����ø������Ҫ�Ķ�ֲ���²�����֮һ��cagA�²�������(Pathogenicity Island, PAI)�����һϵ����ϸ������ת���йصĶ���[3]����Hp�����²����������ܵĻ����в����ˣ�����֤�����Ч���Ͱ�ȫ�ԡ�����Hp�ܴ̼�Tϸ������Fas Ligand (FasL)��ͨ��Fas/FasL����ö���ɱ����ɱ�ˣ���Ҳ����Hp������Tϸ����Ӧ�Խ���Bϸ�����ܵĿ��ܻ��ơ��ڱ��о��У�����Ӧ������Tϸ��ϵģ��������Hp���������ѡ��ԭ��Fas/FasL�鵼Tϸ�������ĵ������ã���Ԥ�������������߱���ʱ�Ŀ����ԡ�
, ��Ĵҽҩ
�����뷽��
һ��Hp�����Tϸ��ϵ
Hp���ʱ���ATCC26695����cagA�²���ͬԴͻ����D-1�������¿���˹��ѧErnst���ڻ���������37�棬��������������������ΪSkirrowѡ������������Һ��Ϊ����������CD4����Tϸ��ϵCem-C7 (Th2����)��Jurkat�������������ռ�������������ģ�����RPMI1640Һ������
����ϸ����������
�����ױ���[4]Ӧ��JAM�����ⶨTϸ��������3H��ǵ�Tϸ����Hp�����������干ͬ����24 h(ϸ��/ϸ������Ϊ1 000��1������1��4ϡ��)���ѽⲢ�ռ�ϸ��DNA����Һ���ⶨ�����ݶ�ʧDNA�ı���������Hp��ɱ���ʡ��ڲⶨHp��ԭ��ɱ������ʱ��Hp��ԭӦ��Ũ�Ⱦ�Ϊ10 ��g/ml��
, ��Ĵҽҩ
����Hp��ԭ
Hp��Ĥ����Hop25��Hop35��Hop38ϵ�ɹ����������ṩ����������ø��ճ����HpaAϵ���дӺ���Hp����ø��������pHP902�ͺ�HpaA��������pHP901�Ĺ��̾��д�����
�ġ���ʽϸ���Ǽ��FasL����
Tϸ����Hp(1��300)���俹ԭ(10 ��g/ml)����24 h���ռ���Ưϴϸ����Ӧ���ÿ���FasL����C-20(Santa Cruz��˾)������FITC�������IgGȾɫ�����տ���Ӧ��������IgG����ʽϸ���Ƿ������Ƚ�Hp��������Ǵ����飬����FasL���������
�塢��������ø���Ƽ���Hpɱ��Tϸ��������
����ϸ�������FasL���ڱ���Һ(��������Һ�е�Ѫ��ɷ�)�еĽ�������ø���Ƽ����⣬��Ϊ������FasL���������Է�����Fas������ϸ����������������Ĥ���FasL������Ӧ�ý�������ø���Ƽ�1,10-phenanthroline(PHE������Sigma��˾)���Fas�鵼��Tϸ����������[5]����ǰ��JAM�����У����Tϸ����Hp����ʱ����Һ����2.5 mmol/L��PHE������ɱ���ʵĸı䡣
, ��Ĵҽҩ
�������ϳ����Ƽ���Hpɱ��Tϸ����FasL�������������
Ϊ̽��Hpɱ��Tϸ���Ƿ���Ҫ�ºϳɵ�FasL���ף�����Ӧ�õ��ϳ����Ƽ�emetine( ����Sigma��˾)����Tϸ��FasL�ϳɼ�Hp�鵼���µ�������[6]��10 mmol/L emetine��Tϸ������45 min����Hp����FasL�����Ӱ�켰�յ�������������
�ߡ�ͳ��ѧ����
���ֵ���(ɱ��)�ʾ���Sigma plot 3.0��ͼ����
��s��ʾ��������student's t���顣P<0.05Ϊ�������������塣���
һ��cagA��Hp��Tϸ�������Ĺ�ϵ
, ��Ĵҽҩ
cagA����Hp�������ǻ�������������������ɱ��Tϸ��Cem-C7��ɱ������43%��53%֮�䣬��CagͬԴͻ����D-1��ɱ����С��10%(P<0.05)����Jurkat��ɱ����ͬ����𣬵�ɱ���ʵ���30%(��1)��
��1 cagA���Լ����Ծ�����Tϸ�������ʱȽ�(%��
��s) Tϸ��ATCC 26695
���
D-1���
ATCC 26695
����
D-1����
, http://www.100md.com
Cem-C7
43��4��
9��4
53��5��
4��2
Jurkat
26��2
6��2
28��4
6��3
����D-1��ȣ�P<0.05
����Hp��ԭ�����Tϸ�������������Ƚ�
, http://www.100md.com
��cagA����Hp�����Ϊ���գ����DZȽ��˼���Hp���浰����Tϸ��Cem-C7���������á��������cagA������ATCC26695�����ճ����HpaA����Ĥ����Hop25��Hop35��Hop38��ɱ���ʷֱ�Ϊ49.0��0.4��18��2��9��4��22��5��75��3��������ø�����á�����Hop38�Ķ�����ǿ��ATCC26695����� Hop25��HpaA�����ý�����Ӧ��Jurkatϸ����ͬ�����֡�
����Hp��������ֶ�Tϸ��FasL����ĵ�������
��ʽϸ�������� Hop38��cagA�����Դ̼�Tϸ������FasL��ͼ1ʾcagA����Hp��ATCC26695�����Tϸ������FasL����cagA����ͻ����D-1�������á��˽����JAM���һ�¡�
�ġ���������ø���Ƽ�������
Ӧ��JAM���������Ƿ����ڽ�������ø���Ƽ�PHE���ڵ�����£�������ATCC26695������������������Cem-C7��Jurkat��ɱ�����þ������(P<0.05)��˵����������ø���Ƽ�������Ĥ���FasL��ɱ�������������ͼ2��
, ��Ĵҽҩ
ͼ1 cagA����Hp���Tϸ��FasL�����Ӱ��
ͼ2 ��������ø���Ƽ���Hp�յ���Tϸ��������Ӱ��
�塢���ϳ����Ƽ�����Hp�鵼��FasL���PTϸ������
10 mmol/L emetine�ܲ����������ϸ���ϳ��µ��ף���ͼ3��ʾ���������µ�Hp26695�鵼��Tϸ��FasL����,ͬʱ����Hp��Cem-C7��ɱ������(ͼ4)��˵��Hp��Tϸ��������Ҫ�ºϳɵ�Fas L���ף�������ֱ��������Tϸ�������Fas���塣
ͼ3 ���ϳ����Ƽ�����Hp�յ���
, http://www.100md.com Tϸ��FasL����
ͼ4 ���ϳ����Ƽ�����Hp��
cem-c7��ɱ������
����
�ڻ����������ԭ�����߷�Ӧ�У�Tϸ����ֱ�Ӳ���ϸ�����߲�����Bϸ���������壬��������ԭ����������Ҳ����Tϸ���鵼�������о����֣�Hp��ȾʱθTϸ����ֳ����Ծ����Hp���������Tϸ����ֳ(����Hp��ԭ���⣬������ø�ɴ̼�Tϸ����ֳ)[7]�����ǻ���Hp��������Tϸ���������书�ܣ��Ӷ�ʹ��Ⱦ����������
�˽�Hp���ܺͱ��濹ԭ��Tϸ��������Ӱ���ܹ�Ԥ������Ϊ����ԭʱ��Ч���Ͱ�ȫ�ԡ�����ø����Tϸ�����������ٽ�����ֳ���Ӷ���������ȫ����Ч������ԭ��cagA�� Hop38��Tϸ�������������ʵ��Ӧ���п������ܴ̼�Tϸ�������㹻��Ч�ĸ���ϸ�����Ӷ����ܻ�ñ�������������������塣HpaA�� Hop 25������������Tϸ�����������ã�������������������á�
, ��Ĵҽҩ
��Ȼ�б���Ӧ��cagA��������С����Ա������ﲻ��cagA����Hp����ĸ�Ⱦ[8]�������о�������һЩ���ֲ���֧��cagA�������ߵ���Чǰ����cagA���Ծ������������֢��/�����صĻ��ߣ���Щ����θ����Ƥ��AP-1��NFkB��ת¼���ӻ�������[9]���伤��ɵ��¶�����֢����������ئ�(IFN-��)������2(IL-2)�������������Ӧ�(TNF-��)��IL-8�ȵı���Ӷ��Ӿ�θճĤ��[10]�����������Tϸ��Fas L���������������������2��NFkB�Ľ��λ�㣬3��AP-1���λ��[11]�����cagA����ͨ������ת¼���Ӷ�����Tϸ��FasL�������������Fas���Ե�θ����Ƥ�����ܴٽ�Tϸ��֮�以���ɱ���Ӿ�����Hp��ԭ����Ӧ�Ժ��������ܡ����о�����cagA���������Ծ����ڼ���FasL��ɱ��Tϸ������ľ����Ϊ������һ�����ṩ���µ�֤�ݡ�
����Fas/ FasL��Hp��ԭ���µ�Tϸ�������е����ã�����T����ϸ���������ֹ��ܱ��ͣ�Th1������IFN-�á�TNF-���ȹ㷺������Hp��Ȼ��Ⱦ��θճĤ�������߱�����Ч�Ķ�����θճĤ��Th2ϸ��Ϊ�����Ҿݱ����������ϸ����Fas/ FasL�鵼ϸ�������������Բ�ͬ����һ�����о�Ӧ�������������Щ�����ѡ��ԭ�Բ�ͬTϸ�����͵����ã��������տ˷�Tϸ�����ܣ���ð�ȫ��Ч��Hp������ԭ��
, ��Ĵҽҩ
������Ŀ��������Ȼ��ѧ����������Ŀ(39700007)
�����
1��D'Elios MM, Manghetti M, Almerigogna F, et al. Different cytokine profile and antigen-specificity repertoire in Helicobacter pylori-specific T cell clones from the antrum of chronic gastritis patients with or without peptic ulcer. Eur J Immunol, 1997,27:1751-1755.
2��Doig P, Trust TJ. Identification of surface-exposed outer membrane antigens of Helicobacter pylori. Infect Immun, 1994,62:4526-4533.
, ��Ĵҽҩ
3��Akopyants NS, Clifton SW, Kersulyte D, et al. Analyses of the cag pathogenicity island of Helicobacter pylori. Mol Microbiol, 1998,28:37-53.
4��Matzinger P. The JAM test. A simple assay for DNA fragmentation and cell death. J Immunol Methods, 1991,145:185.
5��Mitsiades N, Poulaki V, Kotoula V, et al. Fas ligand is present in tumors of the Ewing's sarcoma family and is cleaved into a soluble form by a metalloproteinase. Am J Pathol, 1998,153:1947-1956.
, ��Ĵҽҩ
6��Li JH, Rosen D, Ronen D, et al. The regulation of CD95 ligand expression and function in CTL. J Immunol, 1998,161:3943-3949.
7��Haeberle HA, Kubin M, Bamford KB, et al. Differential stimulation of interleukin-12 (IL-12) and IL-10 by live and killed Helicobacter pylori in vitro and association of IL-12 production with gamma interferon-producing T cells in the human gastric mucosa. Infect Immun, 1997,65:4229-4235.
8��Marchetti M, Rossi M, Giannelli V, et al. Protection against Helicobacter pylori infection in mice by intragastric vaccination with H. pylori antigens is achieved using a non-toxic mutant of E. coli heat-labile enterotoxin (LT) as adjuvant. Vaccine, 1998,16:33-37.
, ��Ĵҽҩ
9��Glocker E, Lange C, Covacci A, et al. Proteins encoded by the cag pathogenicity island of Helicobacter pylori are required for NF-kappa B activation. Infect Immun, 1998,66: 2346-2348.
10��Yasumoto K, Okamoto S, Mukaida N, et al. Tumor necrosis factor and interferon synergistically induce interleukin 8 production in a human gastric cancer cell line through acting concurrently on AP-1 and NF-kappaB-like binding sites of the interleukin 8 gene. J Biol Chem, 1992,267: 22506-22511.
11��Matsui K, Fine A, Ahu B, et al. Identificaiton of two NF-kappa B sites in mouse CD95 ligand (Fas ligand) promoter: Functional analysis in T cell hybridomas. J Immunol, 1998,161: 3469-3473.
(�ո����ڣ�1999-11-25), ��Ĵҽҩ