һ��������ʧͻ��ˮ���ķ��ּ���ϸ��ѧ�о�
���ߣ��ں��� ��ף�� ��־�� ������
��λ�����ݴ�ѧũѧԺũѧϵ�� �������� 225009
�ؼ��ʣ�ˮ����������ʧ
�Ŵ�ѧ��000506
ժҪ����Ʒ���ֻ�422��Ⱥ���з���һ�������꣬��ϸ��ѧ�۲�֤���Ƿ�����������ʧͻ�䣬���ڸ�ֲ��Ļ���ĸϸ���ڼ��������ձ�������ƽ����8.69�������巢����������ʧ��ռ������������72.4%�����ҷ���������ʧ������������ĿΪ12������������ʧ������������Ŀ
�Լ���λϸ���Ķ�����Ƶ�ʣ�ȷ����������ʧ����Ϊ��ȫ������ʧ���졣
��ͼ����ţ�Q343 ���ױ�ʶ�룺A ���±�ţ�0379-4172(2000)05-0418-05
, ��Ĵҽҩ
Isolation and Cytological Identification of a Desynaptic
Variant in Rice
YU HengXiu, CHENG ZhuKuan, GONG ZhiYun, GU MingHong
(Agriculture College Yangzhou University, Yangzhou Jiangsu 225009, China)
Abstract: A sterile variant was found from the population of a japonica rice variety Lunhui 422. Cytological investigation was conducted in meiosis. It is showed that the sterility was caused by desynapsis. As there were 8.69 bivalents in average occurred to be desynaptic, moreover, the most number of the desynaptic bivalents reached 12 at post diakinsis. It can be considered that the desynaptic variation is a
, ��Ĵҽҩ
complete desynapsis found in rice so far.
Key words: rice; desynapsis
������ļ������ѹ�����һ�ֹ����Ժ�ǿ�Ĺ��̣����ֹ����Լȱ�֤������Ⱦɫ����Ŀ�ĺ㶨����ʹ�����Ը�ĸ����Ⱦɫ���ܹ�����ȵؽ��������顣ͬʱ�������ѹ�������һ��ʮ�ָ��ӵĹ��̣��ڸù��������������һЩͻ�䣬�Ӷ��ƻ��������Ĺ����ԡ����йؼ������ѹ����쳣�ı����У�������ʧͻ������Ҫ�ı�������֮һ�����ڼ������Ѵ����ڿɼ���ͬԴȾɫ������ᣬ���ں���I֮ǰ���������������ǰ��ʧ���γ���Ŀ���ȵĵ����塣���ݵ������γ���Ŀ�Ķ��٣����Խ���������Ϊ��ȫ������ʧ�Ͳ���ȫ������ʧ����������ͻ�����ں���I֮ǰ�γɴ��������壬�Ӷ����¼��������γɵ�����Ⱦɫ�岻ƽ�⣬���ںܴ�̶��Ͻ����˱���������ԡ�
������ʧͻ��Ŀǰ�ڴ���[1]��С��[2]������[3]������[4]������[5]���������������в��ٱ�������ˮ�����й�������ʧ��ͻ�����б���[6��15]����Щ����IJ���һ�㶼�����˹��ձ����֯���������������Ȼ״̬�·���������ʧ�����ټ����������о����ֻ�422��Ⱥ��ֲ���з���һ�����꣬��ϸ��ѧ������֤����ֲ�귢����������ʧͻ�䡣�������ر������ǶԸñ������ϸ��ѧ�о������
, http://www.100md.com
1 ���Ϻͷ���
ȡ�����꼰����Ʒ���ֻ�422���������ڵ����룬�Ծƾ�������Ϊ31�Ŀ�ŵ�Ϲ̶�Һ(����0.5% FeCl3)�̶����������Ⱦɫ���о������ڼ��������ڵ�Ⱦɫ����Ϊ��Opton�����������۲���Ӱ��
2 ��������
2.1 ������ʧͻ��ֲ�����̬����
�Ƚ�Ʒ���ֻ�422��������������ʧͻ��������ԣ����Կ��������������ڡ���ߡ�ҶƬ��������ȡ��볤��ÿ�������ȷ���û�����Բ��졣����֮�������Ҫ������ҶƬ�ľ����ij̶��ϣ������ֻ�422ֲ��ҶƬ���ھ����ij̶ȸߣ���ͻ�����ҶƬֻ�������ھ�������ˣ�����ҶƬ�ľ����̶ȣ��������Ե�������������������ʧͻ���ꡣ�ڻ������Լ���ʵ���ϣ���ֲ��������Ϊ���ԣ���I2�KI Ⱦɫ��ͻ����Ļ�����ȫΪ��ܻ���(ͼ��I-A)����Ⱦ������Ϊ0�����������Ⱦ������Ϊ91.5%���ڽ�ʵ���ϣ�ͻ������ȫ��������ʵ��Ϊ0����������Ϊ81.2%��ͻ�����ӱ�����������ֻ�422�Ļ��ۣ��Բ���ʵ��˵���ڻ��۷���������ͬʱ������������˰�����
, ��Ĵҽҩ
2.2 ������ʧͻ��ֲ��ļ���������Ϊ
��������ʧͻ��ֲ��ļ������ѹ��̽��й۲죬��������ڴ����ڣ�ͬԴȾɫ�������������γ�12��������(ͼI-B)���ڴ����ڹ����˫�������ڣ���������������Ľ��棬ͬԴȾɫ������棬�γɽ����(ͼ��I-C)����˵����˫����֮ǰ��ͬԴȾɫ���������ͬʱ���ѷ����������Ⱦɫ����Ľ�������ʱ��δ�е�������γɣ���ˣ�������Ϊ��ʱͬԴȾɫ����������������ġ���˫�������ں��ձ������ڣ��ѿɼ���Ŀ���ȵĵ�������֣��Ƚ��������ͻ��������һʱ�ڵ�Ⱦɫ����̬���Կ���������ͻ�����ʱ���γ���Ŀ���ȵĵ����壬��ʹ�д��������������������Ķ��������(ͼ��I-D)��˵��������ʧ���ձ���֮ǰ�Ѿ��������ڹ�ѧ�����£����ֱ����ѵ��Ա��֡�
���ձ������ڣ�������ʧ������ֵø�Ϊ���ԣ���������Ļ���ĸϸ���ڴ�ʱ���������(ͼ��I-F)����ͬ����ĸϸ������һ����Ŀ�Ķ����巢����������ʧ���۲��������ʱ����������Ϊ8��(ͼ���-G)�����ٵ�Ϊ0��������Ϊ���ж����������ɵ�����(ͼ���-H)��Ϊ��ȷͳ�Ʒ���������ʧ�Ķ�������Ŀ�����ձ������ڵ�����195������ĸϸ�����������1��
, ��Ĵҽҩ
��1 ������ʧͻ�����ڼ��������ձ������ھ��в�ͬ���᷽ʽ�Ļ���ĸϸ���ֲ����
Table 1 Distribution of the PMCs of the desynaptic variant with different synaptic configurations at late diakinesis in meiosis ����
Configuration
ϸ����
No.of cells
�ٷ���
%
����
Configuration
, http://www.100md.com
ϸ����
No.of cells
�ٷ���
%
8��+8��
6
3.08
3��+18��
41
21.03
7��+10��
8
4.10
, ��Ĵҽҩ
2��+20��
38
19.49
6��+12��
11
5.64
1��+22��
26
13.33
5��+14��
22
11.28
24��
, ��Ĵҽҩ
8
4.10
4��+16��
35
17.95
����(Total)
195
100
�ӱ�1���Կ��������л���ĸϸ�����ձ������ھ�����8�����ϵĵ�������֣�����76%���ҵ�ϸ�����ձ������ڳ�����16�����ϵĵ����壬��λϸ���������Ƶ��Ϊ3.31������������ʧ�Ķ�����ռ������������72.4%��˵����ֲ�귢��������ʧͻ���Ƶ�����൱�ߵġ������ڣ������������ڳ�����ϣ�����������Ķ���������(ͼ���-I)�������������ɢ�ڳ��������(ͼ���-J��L)�����ڵ������ں�������ϸ���������κ�һ��������ģ������������˶�ĵ������γɣ���Ȼʹ�ú�������������Ⱦɫ�岻���������ϲ�ƽ�⣬�������ϸ��ѱ�֤����ͬһ����Ⱦɫ��Ϊ
, http://www.100md.com
һ������Ⱦɫ���飬�Ӷ����»��۴������������ڸ�������ʧͻ�������������ֻ�422���۵�������Բ���ʵ��˵���������ӷ���������ͬʱ������������˰�����
3 ����
������ʧͻ����Ϊһ����Ҫ���Ŵ��о����ϣ������о�������ͬԴȾɫ������Ļ����Լ�������������������ȣ����ź�ǿ�������о���Ӧ�ü�ֵ����ˮ�����й�������ʧͻ������һЩ��������������Щͻ���������˳������������ϵ��[6��8]��������Щͻ�������;�������Ǿ������˹��ձ����֯������������У� Kitada�����û�ѧ�ձ��MUN���������11��������ʧͻ����[9]�� Yamguchi�Լ�Ratho�����÷��䴦�������ˮ��������ʧͻ����[10��11]�� Wang�����õ��´����շ���ˮ��������ʧͻ����[12]���趨��������֯���������ɸѡ��һ��ˮ��������ʧͻ����[13]������Щ����ͻ�����;����ͬ���ǣ����о�������ȫ��Ȼ״̬��ɸѡ����������ʧͻ���塣��������ʧͻ��������Ϸ���������ͻ���Ϊ����������ʧ�� Kitada�Ȼ�õ�11��������ʧͻ�����У���9��ͻ�������ֻ�漰1��4�������巢��������ʧ����λ����ĸϸ���У��������Ƶ�ʴ�11.97��10.58��˵����Щ�����������γɶ�����Ϊ����ֻ����������ĸϸ��������������ʧͻ�䡣����2��ͻ�������ֻ�漰6����9�������巢��������ʧ���䵥λϸ��������Ƶ�ʷֱ�Ϊ9.34��6.13���趨��ȴ���֯���������ɸѡ����������ʧͻ���꣬�䷢��������ʧ���Ķ�������ĿΪ5������λϸ��������Ƶ��Ϊ10.40���ӷ���������ʧ���Ķ�������Ŀ�Լ���λϸ���������Ƶ�ʷ�������Щͻ���������������ʧͻ�䡣�����о���õ�������ʧͻ���壬�䷢��������ʧ���Ķ�������ĿΪ12����������Ϊ4������λϸ��������Ƶ��Ϊ3.31������������ʧ�Ķ�����ռȫ���������72.4%��˵���ڸ�������ʧͻ���У�������������ʧ��Ƶ�����൱�ߵģ��������Ϸ����ѽӽ���ȫ������ʧ���������ϣ� Kitada�Ȼ�õ�11��������ʧͻ���壬�䵥λϸ��������Ƶ�����ʵ��֮���������������ء�������Ƶ����ߵ�Ϊ11.97����ʵ��Ϊ82.5%��������Ƶ����͵�Ϊ6.13����ʵ�ʽ�Ϊ1.7%���趨��ȴ�IR54�з��ֵ�ͻ���壬������Ƶ��Ϊ10.40����ʵ��Ϊ2.01%�������о����ֵ�ͻ���壬������72.4%�Ķ����巢����������ʧ����λϸ��������Ƶ�ʽ�Ϊ3.31�����ڴ�����������γɣ��Ӷ����»�����ȫ��������ʵ��Ϊ0��
, ��Ĵҽҩ
��ĿǰΪֹ�����кܶ��о�֤��������ʧͻ���������Ե�������Ƶ�[14��15]����ô���о�����õ�������ʧͻ���Ƿ�Ҳ�����Ե�������ƣ����д���һ��֤ʵ�����⣬��ͻ��������ж����������һ���̶��Ϸ���������ʧ�����������Ʋ⣬���Դ��������Ժ����ѡ��һ��������ϵ�У������ڸ�ֲ����ȫ���������������û�ҩ������;��ʹ�������Ժ�������������ϳ�ȫ������Ĺؼ�������Ʒ���ֻ�422Ϊ����Ʒ�֣�һ���������ܹ����ɣ��ؽ��ı������̵����������ֻ����������Ӧ�õľ����ԣ�ʹ���������̵��Ļ���λ���Ǿ����Ļ���λ��������������Ϊ���߲��ϵ�����ɡ�










, http://www.100md.com
ͼ �� ˵ ��
ͼ��I
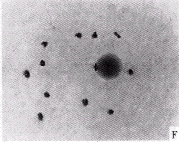
A. ��I2-KIȾɫ���ͻ���껨�ۣ�B. ͻ����Ļ���ĸϸ���ڴ�����ͬԴȾɫ�����������C. ͻ����Ļ���ĸϸ����������˫�������ڳ��������ķֲ棻D. ������Ļ���ϸ�����ձ������ڶ��������̬��E. ͻ����Ļ���ĸϸ�����ձ������ڲ���������ʧ�γɵ����壻F. ������Ļ���ĸϸ�����ձ������ڶ�������̬
ͼ��II
G. ͻ����Ļ���ĸϸ�����ձ������ڲ���������ʧ�γ�8II+8I��H. ͻ����Ļ���ĸϸ�����ձ�������������ʧ�γ�24I��I. ������Ļ���ĸϸ��������I�γ�12II��J. ͻ����Ļ���ĸϸ���������γ�8II+8I��K.ͻ����Ļ���ĸϸ���������γ�1II+22I��L.ͻ����Ļ���ĸϸ��������I�γ�24��������
, ��Ĵҽҩ
Explanation of Plates
Plate I
A. Pollens of the desynaptic variant stained with I2-KI; B. Pachytene chromosomes of the desynaptic variant; C. Chromosomes of the desynaptic variant at early diplotene; D. Chromosomes of the normal plant of LH422 at early diakinesis; E. Chromosomes of the desynaptic variant at early diakinesis; F. Chromosomes of the normal plant of LH422 at late diakinesis
, http://www.100md.com
PlateII
G. Chromosomes of the desynaptic variant at late diakinesis, 8II+8I; H. Chromosomes of the desynaptic variant at late diakinesis,24I��I; . Chromosomes of the normal plant of LH422 at metaphase ,12II; J. Chromosomes of the desynaptic variant at metaphase ,8II+8I; K. Chromosomes of the desynaptic variant at metaphase , 1II+22I; L. Chromosomes of the desynaptic variant at metaphase I,24 I
������Ȼ��ѧ����������Ŀ(��Ŀ���:39700076)
, ��Ĵҽҩ
�����
[1] Enns H, Larter E N. Likage relations of ds, a gene governing chromosome behaviour in barley and its effect on gene recombination. Can. J. Genet. Cytol., 1962, 4:263��266.
[2] Bozzini A, Martini G. Analysis of desynaptic mutants induced in durum wheat (Triticum durum desf.). Caryologia, 1971, 24:307��316.
[3] Ahloowaila B S. Genetic control of post synaptic phase of meiosis in rye grass. Chromosomes Today, 1972, 3:93��103.
, http://www.100md.com
[4] Miller O L. Cytological studies in asynaptic maize. Genetics, 1963, 48:1445��1466.
[5] Krishnaswamy N, Meenakshi K. Abnormal meiosis in grain Sorghum I. Desynapsis. Cytologia, 1957, 22:250��263.
[6] Yasui H, Satoh N, Iwata N. Establishment of a trisomic series in rice by using a desynaptic mutant. Rice Genetic Newsletter, 1989, 6:50��51.
[7] Katayama T. Studies on the progenies of autotriploid and asynaptic rice plant. Japan J. Breed., 1963, 13:83��87.
, http://www.100md.com
[8] Katayama T. Relationships between seed-weight and somatic chromosome number in the progeny of partially asynaptic rice plants: The occurrence of trisomic and monosomic plants. Japan J. Breed., 1966, 16:10��14.
[9] Kitada K, Kurata N, Satoh H et al. Genetic control of meiosis of rice, Oryza sativa L., I. Classification of meiotic mutants induced by MUN and their cytogenetical characteristics. Jpn. J. Genet., 1983, 58:231��240.
, http://www.100md.com [10] Yamaguchi H. Mutations with gamma irradiation on
dormant seeds of a partially asynaptic strain in rice. Japan J. Breed., 1974, 24:81��85.
[11] Ratho S N, Misra R N. Induced desynaptic mutant in rice (Oryza sativa L.). Sci. Cult., 1973, 39:267��269.
[12] Wang S, Yeh P, Lee S et al. Effect of low temperature on desynapsis in rice. Bot. Bull. Acad. Sinica, 1965, 6:197��207.
, ��Ĵҽҩ
[13] �趨��÷��������Ӣ��. ��Դ���̵���ϸ��������������ʧ�����ϸ���Ŵ�ѧ�о�. �Ŵ�ѧ����1988��15(2)��86��88.
[14] Kitada K, Omura T. Genetic control of meiosis of rice, Oryza sativa L. II. Cytological analysis of desynaptic mutants. Jpn. J. Genet., 1983, 58:567��577.
[15] Yasui H, Saiki S, Iwata N. A desynaptic gene ds3(t) located on chromosome 7 in rice. Rice Genetic Newsletter, 1993, 10:80��83.
1999-05-17
1999-11-20, ��Ĵҽҩ
��λ�����ݴ�ѧũѧԺũѧϵ�� �������� 225009
�ؼ��ʣ�ˮ����������ʧ
�Ŵ�ѧ��000506
ժҪ����Ʒ���ֻ�422��Ⱥ���з���һ�������꣬��ϸ��ѧ�۲�֤���Ƿ�����������ʧͻ�䣬���ڸ�ֲ��Ļ���ĸϸ���ڼ��������ձ�������ƽ����8.69�������巢����������ʧ��ռ������������72.4%�����ҷ���������ʧ������������ĿΪ12������������ʧ������������Ŀ
�Լ���λϸ���Ķ�����Ƶ�ʣ�ȷ����������ʧ����Ϊ��ȫ������ʧ���졣
��ͼ����ţ�Q343 ���ױ�ʶ�룺A ���±�ţ�0379-4172(2000)05-0418-05
, ��Ĵҽҩ
Isolation and Cytological Identification of a Desynaptic
Variant in Rice
YU HengXiu, CHENG ZhuKuan, GONG ZhiYun, GU MingHong
(Agriculture College Yangzhou University, Yangzhou Jiangsu 225009, China)
Abstract: A sterile variant was found from the population of a japonica rice variety Lunhui 422. Cytological investigation was conducted in meiosis. It is showed that the sterility was caused by desynapsis. As there were 8.69 bivalents in average occurred to be desynaptic, moreover, the most number of the desynaptic bivalents reached 12 at post diakinsis. It can be considered that the desynaptic variation is a
, ��Ĵҽҩ
complete desynapsis found in rice so far.
Key words: rice; desynapsis
������ļ������ѹ�����һ�ֹ����Ժ�ǿ�Ĺ��̣����ֹ����Լȱ�֤������Ⱦɫ����Ŀ�ĺ㶨����ʹ�����Ը�ĸ����Ⱦɫ���ܹ�����ȵؽ��������顣ͬʱ�������ѹ�������һ��ʮ�ָ��ӵĹ��̣��ڸù��������������һЩͻ�䣬�Ӷ��ƻ��������Ĺ����ԡ����йؼ������ѹ����쳣�ı����У�������ʧͻ������Ҫ�ı�������֮һ�����ڼ������Ѵ����ڿɼ���ͬԴȾɫ������ᣬ���ں���I֮ǰ���������������ǰ��ʧ���γ���Ŀ���ȵĵ����塣���ݵ������γ���Ŀ�Ķ��٣����Խ���������Ϊ��ȫ������ʧ�Ͳ���ȫ������ʧ����������ͻ�����ں���I֮ǰ�γɴ��������壬�Ӷ����¼��������γɵ�����Ⱦɫ�岻ƽ�⣬���ںܴ�̶��Ͻ����˱���������ԡ�
������ʧͻ��Ŀǰ�ڴ���[1]��С��[2]������[3]������[4]������[5]���������������в��ٱ�������ˮ�����й�������ʧ��ͻ�����б���[6��15]����Щ����IJ���һ�㶼�����˹��ձ����֯���������������Ȼ״̬�·���������ʧ�����ټ����������о����ֻ�422��Ⱥ��ֲ���з���һ�����꣬��ϸ��ѧ������֤����ֲ�귢����������ʧͻ�䡣�������ر������ǶԸñ������ϸ��ѧ�о������
, http://www.100md.com
1 ���Ϻͷ���
ȡ�����꼰����Ʒ���ֻ�422���������ڵ����룬�Ծƾ�������Ϊ31�Ŀ�ŵ�Ϲ̶�Һ(����0.5% FeCl3)�̶����������Ⱦɫ���о������ڼ��������ڵ�Ⱦɫ����Ϊ��Opton�����������۲���Ӱ��
2 ��������
2.1 ������ʧͻ��ֲ�����̬����
�Ƚ�Ʒ���ֻ�422��������������ʧͻ��������ԣ����Կ��������������ڡ���ߡ�ҶƬ��������ȡ��볤��ÿ�������ȷ���û�����Բ��졣����֮�������Ҫ������ҶƬ�ľ����ij̶��ϣ������ֻ�422ֲ��ҶƬ���ھ����ij̶ȸߣ���ͻ�����ҶƬֻ�������ھ�������ˣ�����ҶƬ�ľ����̶ȣ��������Ե�������������������ʧͻ���ꡣ�ڻ������Լ���ʵ���ϣ���ֲ��������Ϊ���ԣ���I2�KI Ⱦɫ��ͻ����Ļ�����ȫΪ��ܻ���(ͼ��I-A)����Ⱦ������Ϊ0�����������Ⱦ������Ϊ91.5%���ڽ�ʵ���ϣ�ͻ������ȫ��������ʵ��Ϊ0����������Ϊ81.2%��ͻ�����ӱ�����������ֻ�422�Ļ��ۣ��Բ���ʵ��˵���ڻ��۷���������ͬʱ������������˰�����
, ��Ĵҽҩ
2.2 ������ʧͻ��ֲ��ļ���������Ϊ
��������ʧͻ��ֲ��ļ������ѹ��̽��й۲죬��������ڴ����ڣ�ͬԴȾɫ�������������γ�12��������(ͼI-B)���ڴ����ڹ����˫�������ڣ���������������Ľ��棬ͬԴȾɫ������棬�γɽ����(ͼ��I-C)����˵����˫����֮ǰ��ͬԴȾɫ���������ͬʱ���ѷ����������Ⱦɫ����Ľ�������ʱ��δ�е�������γɣ���ˣ�������Ϊ��ʱͬԴȾɫ����������������ġ���˫�������ں��ձ������ڣ��ѿɼ���Ŀ���ȵĵ�������֣��Ƚ��������ͻ��������һʱ�ڵ�Ⱦɫ����̬���Կ���������ͻ�����ʱ���γ���Ŀ���ȵĵ����壬��ʹ�д��������������������Ķ��������(ͼ��I-D)��˵��������ʧ���ձ���֮ǰ�Ѿ��������ڹ�ѧ�����£����ֱ����ѵ��Ա��֡�
���ձ������ڣ�������ʧ������ֵø�Ϊ���ԣ���������Ļ���ĸϸ���ڴ�ʱ���������(ͼ��I-F)����ͬ����ĸϸ������һ����Ŀ�Ķ����巢����������ʧ���۲��������ʱ����������Ϊ8��(ͼ���-G)�����ٵ�Ϊ0��������Ϊ���ж����������ɵ�����(ͼ���-H)��Ϊ��ȷͳ�Ʒ���������ʧ�Ķ�������Ŀ�����ձ������ڵ�����195������ĸϸ�����������1��
, ��Ĵҽҩ
��1 ������ʧͻ�����ڼ��������ձ������ھ��в�ͬ���᷽ʽ�Ļ���ĸϸ���ֲ����
Table 1 Distribution of the PMCs of the desynaptic variant with different synaptic configurations at late diakinesis in meiosis ����
Configuration
ϸ����
No.of cells
�ٷ���
%
����
Configuration
, http://www.100md.com
ϸ����
No.of cells
�ٷ���
%
8��+8��
6
3.08
3��+18��
41
21.03
7��+10��
8
4.10
, ��Ĵҽҩ
2��+20��
38
19.49
6��+12��
11
5.64
1��+22��
26
13.33
5��+14��
22
11.28
24��
, ��Ĵҽҩ
8
4.10
4��+16��
35
17.95
����(Total)
195
100
�ӱ�1���Կ��������л���ĸϸ�����ձ������ھ�����8�����ϵĵ�������֣�����76%���ҵ�ϸ�����ձ������ڳ�����16�����ϵĵ����壬��λϸ���������Ƶ��Ϊ3.31������������ʧ�Ķ�����ռ������������72.4%��˵����ֲ�귢��������ʧͻ���Ƶ�����൱�ߵġ������ڣ������������ڳ�����ϣ�����������Ķ���������(ͼ���-I)�������������ɢ�ڳ��������(ͼ���-J��L)�����ڵ������ں�������ϸ���������κ�һ��������ģ������������˶�ĵ������γɣ���Ȼʹ�ú�������������Ⱦɫ�岻���������ϲ�ƽ�⣬�������ϸ��ѱ�֤����ͬһ����Ⱦɫ��Ϊ
, http://www.100md.com
һ������Ⱦɫ���飬�Ӷ����»��۴������������ڸ�������ʧͻ�������������ֻ�422���۵�������Բ���ʵ��˵���������ӷ���������ͬʱ������������˰�����
3 ����
������ʧͻ����Ϊһ����Ҫ���Ŵ��о����ϣ������о�������ͬԴȾɫ������Ļ����Լ�������������������ȣ����ź�ǿ�������о���Ӧ�ü�ֵ����ˮ�����й�������ʧͻ������һЩ��������������Щͻ���������˳������������ϵ��[6��8]��������Щͻ�������;�������Ǿ������˹��ձ����֯������������У� Kitada�����û�ѧ�ձ��MUN���������11��������ʧͻ����[9]�� Yamguchi�Լ�Ratho�����÷��䴦�������ˮ��������ʧͻ����[10��11]�� Wang�����õ��´����շ���ˮ��������ʧͻ����[12]���趨��������֯���������ɸѡ��һ��ˮ��������ʧͻ����[13]������Щ����ͻ�����;����ͬ���ǣ����о�������ȫ��Ȼ״̬��ɸѡ����������ʧͻ���塣��������ʧͻ��������Ϸ���������ͻ���Ϊ����������ʧ�� Kitada�Ȼ�õ�11��������ʧͻ�����У���9��ͻ�������ֻ�漰1��4�������巢��������ʧ����λ����ĸϸ���У��������Ƶ�ʴ�11.97��10.58��˵����Щ�����������γɶ�����Ϊ����ֻ����������ĸϸ��������������ʧͻ�䡣����2��ͻ�������ֻ�漰6����9�������巢��������ʧ���䵥λϸ��������Ƶ�ʷֱ�Ϊ9.34��6.13���趨��ȴ���֯���������ɸѡ����������ʧͻ���꣬�䷢��������ʧ���Ķ�������ĿΪ5������λϸ��������Ƶ��Ϊ10.40���ӷ���������ʧ���Ķ�������Ŀ�Լ���λϸ���������Ƶ�ʷ�������Щͻ���������������ʧͻ�䡣�����о���õ�������ʧͻ���壬�䷢��������ʧ���Ķ�������ĿΪ12����������Ϊ4������λϸ��������Ƶ��Ϊ3.31������������ʧ�Ķ�����ռȫ���������72.4%��˵���ڸ�������ʧͻ���У�������������ʧ��Ƶ�����൱�ߵģ��������Ϸ����ѽӽ���ȫ������ʧ���������ϣ� Kitada�Ȼ�õ�11��������ʧͻ���壬�䵥λϸ��������Ƶ�����ʵ��֮���������������ء�������Ƶ����ߵ�Ϊ11.97����ʵ��Ϊ82.5%��������Ƶ����͵�Ϊ6.13����ʵ�ʽ�Ϊ1.7%���趨��ȴ�IR54�з��ֵ�ͻ���壬������Ƶ��Ϊ10.40����ʵ��Ϊ2.01%�������о����ֵ�ͻ���壬������72.4%�Ķ����巢����������ʧ����λϸ��������Ƶ�ʽ�Ϊ3.31�����ڴ�����������γɣ��Ӷ����»�����ȫ��������ʵ��Ϊ0��
, ��Ĵҽҩ
��ĿǰΪֹ�����кܶ��о�֤��������ʧͻ���������Ե�������Ƶ�[14��15]����ô���о�����õ�������ʧͻ���Ƿ�Ҳ�����Ե�������ƣ����д���һ��֤ʵ�����⣬��ͻ��������ж����������һ���̶��Ϸ���������ʧ�����������Ʋ⣬���Դ��������Ժ����ѡ��һ��������ϵ�У������ڸ�ֲ����ȫ���������������û�ҩ������;��ʹ�������Ժ�������������ϳ�ȫ������Ĺؼ�������Ʒ���ֻ�422Ϊ����Ʒ�֣�һ���������ܹ����ɣ��ؽ��ı������̵����������ֻ����������Ӧ�õľ����ԣ�ʹ���������̵��Ļ���λ���Ǿ����Ļ���λ��������������Ϊ���߲��ϵ�����ɡ�
, http://www.100md.com
ͼ �� ˵ ��
ͼ��I
A. ��I2-KIȾɫ���ͻ���껨�ۣ�B. ͻ����Ļ���ĸϸ���ڴ�����ͬԴȾɫ�����������C. ͻ����Ļ���ĸϸ����������˫�������ڳ��������ķֲ棻D. ������Ļ���ϸ�����ձ������ڶ��������̬��E. ͻ����Ļ���ĸϸ�����ձ������ڲ���������ʧ�γɵ����壻F. ������Ļ���ĸϸ�����ձ������ڶ�������̬
ͼ��II
G. ͻ����Ļ���ĸϸ�����ձ������ڲ���������ʧ�γ�8II+8I��H. ͻ����Ļ���ĸϸ�����ձ�������������ʧ�γ�24I��I. ������Ļ���ĸϸ��������I�γ�12II��J. ͻ����Ļ���ĸϸ���������γ�8II+8I��K.ͻ����Ļ���ĸϸ���������γ�1II+22I��L.ͻ����Ļ���ĸϸ��������I�γ�24��������
, ��Ĵҽҩ
Explanation of Plates
Plate I
A. Pollens of the desynaptic variant stained with I2-KI; B. Pachytene chromosomes of the desynaptic variant; C. Chromosomes of the desynaptic variant at early diplotene; D. Chromosomes of the normal plant of LH422 at early diakinesis; E. Chromosomes of the desynaptic variant at early diakinesis; F. Chromosomes of the normal plant of LH422 at late diakinesis
, http://www.100md.com
PlateII
G. Chromosomes of the desynaptic variant at late diakinesis, 8II+8I; H. Chromosomes of the desynaptic variant at late diakinesis,24I��I; . Chromosomes of the normal plant of LH422 at metaphase ,12II; J. Chromosomes of the desynaptic variant at metaphase ,8II+8I; K. Chromosomes of the desynaptic variant at metaphase , 1II+22I; L. Chromosomes of the desynaptic variant at metaphase I,24 I
������Ȼ��ѧ����������Ŀ(��Ŀ���:39700076)
, ��Ĵҽҩ
�����
[1] Enns H, Larter E N. Likage relations of ds, a gene governing chromosome behaviour in barley and its effect on gene recombination. Can. J. Genet. Cytol., 1962, 4:263��266.
[2] Bozzini A, Martini G. Analysis of desynaptic mutants induced in durum wheat (Triticum durum desf.). Caryologia, 1971, 24:307��316.
[3] Ahloowaila B S. Genetic control of post synaptic phase of meiosis in rye grass. Chromosomes Today, 1972, 3:93��103.
, http://www.100md.com
[4] Miller O L. Cytological studies in asynaptic maize. Genetics, 1963, 48:1445��1466.
[5] Krishnaswamy N, Meenakshi K. Abnormal meiosis in grain Sorghum I. Desynapsis. Cytologia, 1957, 22:250��263.
[6] Yasui H, Satoh N, Iwata N. Establishment of a trisomic series in rice by using a desynaptic mutant. Rice Genetic Newsletter, 1989, 6:50��51.
[7] Katayama T. Studies on the progenies of autotriploid and asynaptic rice plant. Japan J. Breed., 1963, 13:83��87.
, http://www.100md.com
[8] Katayama T. Relationships between seed-weight and somatic chromosome number in the progeny of partially asynaptic rice plants: The occurrence of trisomic and monosomic plants. Japan J. Breed., 1966, 16:10��14.
[9] Kitada K, Kurata N, Satoh H et al. Genetic control of meiosis of rice, Oryza sativa L., I. Classification of meiotic mutants induced by MUN and their cytogenetical characteristics. Jpn. J. Genet., 1983, 58:231��240.
, http://www.100md.com [10] Yamaguchi H. Mutations with gamma irradiation on
dormant seeds of a partially asynaptic strain in rice. Japan J. Breed., 1974, 24:81��85.
[11] Ratho S N, Misra R N. Induced desynaptic mutant in rice (Oryza sativa L.). Sci. Cult., 1973, 39:267��269.
[12] Wang S, Yeh P, Lee S et al. Effect of low temperature on desynapsis in rice. Bot. Bull. Acad. Sinica, 1965, 6:197��207.
, ��Ĵҽҩ
[13] �趨��÷��������Ӣ��. ��Դ���̵���ϸ��������������ʧ�����ϸ���Ŵ�ѧ�о�. �Ŵ�ѧ����1988��15(2)��86��88.
[14] Kitada K, Omura T. Genetic control of meiosis of rice, Oryza sativa L. II. Cytological analysis of desynaptic mutants. Jpn. J. Genet., 1983, 58:567��577.
[15] Yasui H, Saiki S, Iwata N. A desynaptic gene ds3(t) located on chromosome 7 in rice. Rice Genetic Newsletter, 1993, 10:80��83.
1999-05-17
1999-11-20, ��Ĵҽҩ