炎症反应Toll信号传导通路
作者:王伯瑶 黄宁 吴琦
单位:(华西医科大学感染免疫研究室 病理生理教研室, 四川 成都 610041)
关键词:炎症;信号传递;基因
中国病理生理杂志000625 [中图分类号] R364.5 [文献标识码] A
[文章编号] 1000-4718(2000)06-0567-06
Toll receptor signaling pathway in inflammation
WANG Bo-yao, HUANG Ning, WU Qi
(Research Unit of Infection & Immunity, Department of Pathophysiology, West China University of Medical Sciences, Chengdu 610041, China)
, 百拇医药
【A Review】 Toll signaling pathway may play a curcial role in induction of inflammation-associated gene activation. Originally, the Toll/spaetzle/Cactus-Dorsal signaling pathway is established in the Drosophila embryonic development. Recently, the Toll signaling pathway in adult Drosophila has been established in the induction of antimicrobial peptide expression. Five human Toll-like receptor genes (hTlr l-5) and one mouse Toll-like receptor gene (mTlr-4) have been isolated. Toll and Toll-like receptor genes encoded molecules are transmembrane proteins with an extracellular leucine repeat domain and a cytoplasmic domain homologous to IL-1 receptors. The intracellular signaling cascade involves Tube, Pelle, and Cactus-Dorsal complex in Drosophila, and MyD88, IRAK, TRAF 6, NIK, αβ-I κB kinase, and I κB -NFκB complex in mammals. Dorsal and NFκB are transcription factors, while Cactus and IκB are their inhibitors. When the inhibitors phosphorylated, the nuclear factors are released and move into nucleus, leading to immune gene activation. It has been shown from in vitro system that Tlr-4 mediated LPS signaling in human monocytes for expression of IL-1, IL-6, IL-8, and costimulator B7-1 which provides second signal for T cell response. Tlr-2 can also mediate LPS signaling in human monocytes, leading to the production of proinflammatory mediators. Microbial lipoproteins are potent stimulators of IL-12 production through Tlr-2 signaling by human macrophages, and can stimulate Tlr 2-dependent transcription of inducible nitric oxide synthase and the production of nitric oxide, a powerful microbicidal pathway. Findings of a point mutation of Tlr-4 in LPS tolerant C3H/HeJ mouse strain and a deletion of Tlr-4 in LPS resistant C57BL/10ScCr mice provide an in vivo evidence strongly supporting the crucial role of Tlrs in LPS mediated inflammation. It is proposed that targeting Tlrs would develop new remedies for treatment of inflammatory disorders and for immunotherapy of mucosal infections and cancer, etc.
, 百拇医药
[MeSH] Inflammation; Signal transduction; Genes
炎症反应是机体天然抵抗微生物侵袭的重要武器[1],因此从免疫学角度讲又称之为天然免疫。补体系统、凝血系统、急性期反应蛋白、吞噬细胞、内皮细胞和上皮细胞等构成这个反应系统的主要元件。天然免疫在发生学上是有机体最古老的抗感染防御机制,它使用胚胎细胞编码的形态识别受体(pattern recognition receptors, PRRs) 识别微生物的保守的分子成分,如LPS、胞壁脂蛋白、甘露糖和RNA病毒双链RNA等,产生抗微生物及其毒性因子的免疫反应[2]。这种分子形态识别受体从分布上可分为细胞性受体和体液性受体;从功能上可分为循环血中的体液性受体和在细胞膜上表达的内吞受体以及信号受体等。体液性受体如甘露糖结合蛋白、C反应蛋白、血清淀粉样蛋白等介导补体系统的激活和调理吞噬作用,而脂多糖结合蛋白(LBP)识别LPS,起浓集血中LPS并传递给CD14的作用,它们均属于由肝细胞合成的急性期反应蛋白;内吞受体如巨噬细胞甘露糖受体、清道夫受体和补体受体等介导吞噬作用;信号受体如CD14和Toll在细胞介导的炎症反应中起信号传导作用。天然免疫细胞识别病原体相关分子形态(pathogen associated molecular patterns, PAMs)或蛋白水解信号,一方面诱导抗菌肽的合成,直接杀伤侵入的病原微生物;另一方面诱导内源性信号的表达,如炎症预激细胞因子(TNF、IL-1等)、趋化细胞因子(IL-8、MCP、MIP等)、粘附分子、共刺激因子(B7、IL-12等)以及一氧化氮合酶(产生一氧化氮)等,以便更有效的动员机体的防御力量来抵抗病原微生物的侵入。人们已熟知髓性白细胞膜CD14介导LPS信号胞内传导而使NF-κB激活,以诱导上述内源性信号分子基因的表达。然而,CD14分子缺乏胞浆区段,不能直接向细胞内传导LPS信号,尚需其它分子起信号传导作用。新近发现的免疫调节基因产物Toll蛋白可能是其信号向胞内传导的门户蛋白(gateway),本文概要综述其研究进展。
, 百拇医药
一、从果蝇发育调节的研究发现免疫信号受体基因Toll
在果蝇胚胎的发育成形过程中最重要阶段是其受精卵向前后轴和背腹轴分化[3]。用遗传分析法在80年代发现了母体基因座Toll调控果蝇胚基细胞向背腹轴方向分化发育,形成胚胎的外胚层和中胚层[4,5]。以后用遗传学、分子生物学和生物化学方法确立了Toll基因产物Toll蛋白是分布在胚胎细胞上的信号受体,并确立了信号传导通路Spaetzle/Toll/Cactus-Dorsal[5]。Spaetzle基因编码47~60 kD蛋白质,分泌到卵黄周围间隙(卵黄膜和胚胎浆膜之间充满液体的空间),在4个丝氨酸蛋白酶家族成员级联反应作用下,最终使其水解产生23 kD的Spaetzle活性蛋白,作为Toll受体的配体而激活Toll蛋白,导致细胞内信号传导级联反应,即激活的Toll蛋白通过辅助因子Tube而使丝氨酸/苏氨酸蛋白激酶Pelle激活,Cactus的磷酸化和降解使转录因子Dorsal从无活性的胞浆蛋白复合物Cactus-Dorsal中释放出来,并移位到核内,激活相应的形态发生基因,导致果蝇胚胎细胞向背腹轴方向分化[6]。
, 百拇医药
在80年代初对果蝇抗感染机制的研究发现了抗菌肽,并被证明是果蝇抗感染的主要介质[7]。抗菌肽并不在果蝇体内固有表达,仅在感染性损伤或注射微生物成分和LPS时才在果蝇肥体细胞(在功能上相当于哺乳动物的肝细胞)和/或上皮细胞被诱导表达。现已知被诱导产生的抗菌肽分子至少有7种,即cecropins\, diptericin\, drosocin\, attacin\, 防御素和metchnikowin, 它们主要起抗细菌作用;而drosomycin具强力抗霉菌活性。所有这些抗菌肽基因的上游序列均含转录因子Dorsal结合位点,新近的实验研究已显示果蝇胚胎背腹轴成形级联反应调节基因Spaetzle/Toll/Cactus调控成年果蝇抗菌肽的表达[8]。Dorsal属于Rel蛋白家族成员,该蛋白家族是可迅速被诱导激活的基因的转录因子,在果蝇包括Dorsal和Dif蛋白;在哺乳动物是免疫和炎症反应效应基因转录激活因子NF-κB[9]。通过Database检索和分子同源性比较,发现IL-1诱导哺乳动物炎症反应中NF-κB激活的级联反应与果蝇Spaetzle/Toll/Cactus-Dorsal通路相似, 从而使学者们对Toll受体信号通路在哺乳动物天然免疫和炎症反应中的作用产生极大兴趣。现已克隆出5个人Toll样受体基因,称为Tlrs1-5(Toll-like receptors-1-5)[10],Tlrs又称hTolls。并相继在多个国际著名实验室和医药公司对其结构与功能进行热火朝天的研究[1]。
, 百拇医药
二、Toll蛋白的结构与功能的研究进展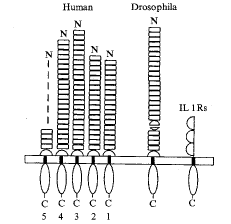
Fig 1 Schematic structure of Toll and Toll-like receptors (from Rock FL, et al.Proc Natl Acad Sci USA. 1998, 95:588 with some modification)
图1 Toll及Toll样受体的基本结构
如图1所示,Toll属于I型跨膜受体蛋白,其细胞外区段(extracellular domain)由富含亮氨酸的重复序列和半胱氨酸结(半胱氨酸残基形成链内二硫键袢)所组成,果蝇Toll含2个半胱氨酸结,使其细胞外区段被第二个半胱氨酸结分隔成2个节段,而人Tlrs的细胞外区段仅含1个半胱氨酸结,紧邻细胞外膜;其细胞内胞浆区段(cytoplasmic domain)与哺乳动物IL-1受体的胞浆区段具同源性,果蝇Toll蛋白的胞浆区段的氨基酸序列与哺乳动物IL-1受体分子胞浆序列的同一性为26%,其相似性为43%[10,11]。然而,Toll蛋白的细胞外区段显然与IL-1受体相异,后者在结构上属Ig超家族。细胞膜CD14是一富含亮氨酸重复序列的糖基磷脂酰肌醇锚定蛋白,虽然与Toll蛋白细胞外区段在结构上具某种相似性,但缺乏胞浆区段,不能向细胞内传递信息。因此,Toll蛋白应为一类新的信号受体蛋白。
, 百拇医药
现克隆出的人Tlrs全长cDNAs分别为:Tlr-1, 2 366 bp,其开放读框编码786个氨基酸肽链;Tlr-2, 2 600 bp,其开放读框编码784个氨基酸肽链;Tlr-3, 3 029 bp, 其开放读框编码904个氨基酸肽链;Tlr-4, 3 811 bp,其开放读框编码879个氨基酸肽链;Tlr-5, 1 275 bp,其开放读框编码370个氨基酸肽链。以Tlrs cDNAs作探针进行荧光素标记原位杂交(FISH)染色体定位分析显示Tlr1-3基因定位于4号染色体上,Tlr4基因定位于9号染色体上,Tlr5基因定位于1号染色体上[10]。
Fig 2 Construction of an autologously activated Tlr4 chimaric gene (from Medzhitov R, et al. Nature, 1997, 388:394. AFR and CFR represent the amino-terminal flanking and carboxy-terminal cysteine-rich regions. LRR, leucine repeat region. TM, transmembrane region)
, 百拇医药
图2 自发激活型Tlr4嵌合基因的构建
关于Tlrs各成员的信号传递功能,现有报道显示Tlr-4跨膜段和胞浆区段与小鼠CD4胞外区段的编码基因片段构建的自身激活型嵌合基因,转染人髓系单核细胞系THP-1,无需配体即能激活NF-κB和表达炎症细胞因子IL-1、IL-6、IL-8及其共刺激分子B7-1(该分子为T细胞激活所必需的第二信号)[12]。该重组突变基因的构建见示意图2。
Tlr-2编码基因转染不表达Tlrs和CD14的人胚肾细胞系293,能使其接受LPS-LBP复合物刺激信号,诱导NF-κB激活和内皮选择素(E-selectin)基因的转录,而暂时共转染膜CD14编码基因的293细胞提高了对LPS反应敏感性和强度[13,14]。如图3所示,Tlr-2跨膜段和胞浆区段编码基因与CD4细胞外区段编码基因构建的CD4-Tlr-2嵌合基因,以及删除Tlr-2胞浆区段13位或141位氨基酸以后的编码序列而构建的缺失突变基因Tlr2-1和Tlr2-2,转染293细胞,对LPS反应性均丧失,前者表明Tlr-2细胞外区段为其识别LPS所必需的元件,后者提示其羧基端氨基酸序列为细胞内信号传导所必需,与IL-1受体膜内区段信号传导所必需的氨基酸同一性很高,见图3。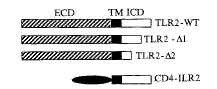
, 百拇医药
Fig 3 Construction of deletion mutants of Tlr-2 gene (from Yang R-B, et al. Nature 1998, 395:284.ECD, extracellular domain. ICD, intracellular domain. TM, transmembrane region)
图3 Tlr-2缺失突变基因的构建
由于人胚肾293细胞系既不表达CD14和Tlrs也不表达炎症细胞因子,已作为一个体外模式系统用于人Tlrs基因的功能研究。最新研究报道[15]显示细菌胞壁脂蛋白,如结核杆菌19 kD脂蛋白、burgdorferi包柔氏螺旋体脂蛋白(OspA)和梅毒螺旋体47 kD脂蛋白等,能刺激转染了Tlr-2编码基因的293细胞系的IL-12基因的转录激活,共转染CD14编码基因加强这种转录激活作用;Tlr-2胞浆区段缺失突变基因转染RAW264.7巨噬细胞系,抑制细菌脂蛋白介导的IL-12和一氧化氮合酶基因的转录激活。亦有报道细菌脂蛋白能介导转染了Tlr-2编码基因的293细胞的凋亡,提示Tlrs可能在炎症反应的负相调节机制中发挥作用[16]。
, 百拇医药
三、感染性炎症反应的Toll信号转导通路[12, 17, 18]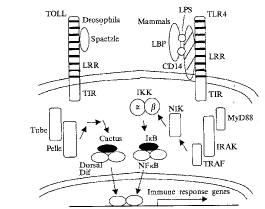
Fig 4 Toll and Toll-like receptor-mediated intracellular signaling (From Hoffman JA, et al. Science, 1999, 284:1313 with a little modification)
图4 Toll及Toll样受体介导的细胞内信号传递
如图4所示,左边代表Toll信号通路诱导成年果蝇抗霉菌基因表达抗霉菌多肽drosomycin,右边代表LPS-LBP-CD14复合物与人Tlr-4结合而激活共刺激因子B7基因的胞内信号传导通路。当成年果蝇受到霉菌感染时,通过其尚未明了的体液性形态识别受体与病原体相关形态分子结合而触发淋巴液中一系列酶解反应,产生激活型Spaetzle而激活Toll蛋白,引起胞内信号传导级联反应,这个反应涉及Tube蛋白、丝氨酸/苏氨酸激酶Pelle,最终使Cactus磷酸化而降解,释放转录因子Dorsal,激活的Dorsal向细胞核移位,与抗霉菌基因增强子序列中的顺式作用元件结合而使其激活,合成抗霉菌多肽。显然该通路与调控果蝇胚胎早期发育背腹轴细胞分化的Toll信号传导通路一致。LPS-LBP-CD14 复合物与人单核吞噬细胞Tlr-4结合触发胞内级联信号传导反应,首先Tlr-4胞浆区段与一个叫做衔接子(adapter)的蛋白MyD88相互作用,依次激活IL-1受体相关激酶(IRAK)、肿瘤坏死因子受体相关因子-6(TRAF6)、NF-κΒ诱导激酶(NIK)和αβ-IκB激酶,最终使IκB磷酸化而降解,激活的NF-κB移位入核内,启动免疫反应基因转录激活。在哺乳动物Toll介导的胞内信号传导所涉及的重要分子无论在结构上或在功能上均与成年果蝇Toll介导的免疫信号胞内传导通路(如NF-κB与Dorsal; IκB与Cactus; IRAK与Pelle等)具相似性,说明Toll介导的免疫和炎症信号传导通路起源古老和保守。
, http://www.100md.com
四、Toll基因突变与内毒素耐受性[19~21]
依据人Tlr-4基因序列设计引物,应用逆转录-聚合酶链反应(RT-PCR)技术已分离出小鼠Tlr-4 cDNA,其编码的氨基酸序列与人Tlr-4具高度同源性(同一性为67%, 相似性为79%),而胞浆区段的同源性比胞外区段高(前者为89%,后者为68%)。
人们已熟知细菌LPS诱发感染宿主强烈的全身炎症反应。在小鼠,这一反应的遗传调节定位于4号染色体Lps基因座,存在2个等位基因,即LPS低反应基因(Lpsd)和LPS反应基因(Lpsn)。C3H/HeJ和C57BL/10ScCr是Lpsd-d纯合子小鼠系,它们对LPS的攻击不发生全身炎症反应,即对LPS具耐受性,然而其分子遗传机制尚未明了。最新研究报道用遗传学和限制性内切酶作图技术确定Lps基因座图距为0.9厘摩(cM),核苷酸序列跨距为1.7 Mb,包含3个转录单位,其中之一为Tlr-4基因。C3H/HeJ小鼠系Tlr-4基因编码区发生了点突变,即712位上编码脯氨酸的密码子由编码组氨酸的密码子替换了,其结果是合成的蛋白分子712位上(位于Tlr-4肽链的胞浆区段)的脯氨酸变成了碱性组氨酸,推测这一氨基酸的突变可能使Tlr-4胞浆区段构型发生改变而不能与下游分子发生相互作用,致使LPS通过Tlr-4蛋白介导的胞内信号传导被中断。而在C57BL/10ScCr小鼠系Lps基因座中却表现Tlr-4基因的缺失。这些研究结果提示Tlr-4结构与功能的变化与机体对内毒素的耐受性密切相关,另一方面从整体水平上提供了证据表明Tlrs信号受体在介导炎症刺激信号的胞内传导中起重要作用。
, 百拇医药
五、小结与展望
在果蝇胚胎发育研究中发现其背腹成形受Toll基因的调控。新近发现该基因亦是病原微生物诱导成年果蝇产生抗菌肽的信号传导通路的门户蛋白基因,从人亦克隆出5个Toll样蛋白家族成员,从小鼠克隆出一个与人Tlr-4具高度同源性的基因。体外和体内实验研究均显示Tlr-4介导内毒素信号胞内传导,激活免疫和炎症相关基因,产生相应的细胞因子或其它介质。在体外实验系统,亦显示Tlr-2可介导内毒素和细菌脂蛋白信号的胞内传导。这些实验结果提供了一个有力证据表明炎症反应与生物的发育、进化发展密切相关,即发育调控基因的趋异(divergent evolution)进化分化出免疫调节基因,以抵抗病原微生物的侵袭,保证复细胞生物的发育和进化。天然免疫系统起源古老,其调控分子相当保守。因此,研究Toll及其相关基因结构与功能可能是揭示哺乳动物和人炎症反应的本质特征的一个突破口。
无论是感染性炎症反应或非感染性慢性炎症所导致的组织器官损伤和疾病状态,如全身炎症反应综合征所致的休克及多器官功能衰竭、进行性毁损性青少年牙周炎、动脉粥样硬化等,均只在一定的易感人群中发生,提示可能有其遗传背景,C3H/HeJ小鼠系对高脂食物诱发动脉粥样硬化具耐受性[22]已提供证据支持这种假设。在对内毒素耐受小鼠系C3H/HeJ和C57BL/10ScCr上的分子遗传研究显示Tlr-4基因的突变是其内毒素耐受的原因,这为揭示上述炎症疾病的分子遗传基础的研究提供了一条令人鼓舞的途径。
, http://www.100md.com
从信号受体Toll蛋白基因的研究显示,炎症反应是低等动物发育过程中分化出的抗微生物感染的天然防御机制。在低等的无脊椎动物如昆虫,其生物学效应主要是产生抗菌肽,以行使抗感染作用;随进化发展,到了哺乳动物和人,炎症反应分化发展得相当复杂了。有体液反应系统和细胞介导系统;在细胞介导系统中,有被诱导产生的各种炎症细胞因子、其它小分子炎症介质以及抗菌肽;其反应或强或弱;既可产生保护效应又可发生损伤后果。深入研究古老的Toll及其相关基因的结构和功能可能对这一复杂反应理出一定头绪。比如,抗菌肽的产生和对细胞介导反应的正相与负相调节起作用的各种内源性信号的表达可能由不同的Toll样信号受体介导; 在体液反应系统的级联反应酶解产物中可能存在Spaetzle相关分子,这方面的研究有可能把体液反应系统与细胞介导系统联系起来,且可推进和加深对非感染性炎症疾病如自身免疫疾病和动脉粥样硬化发病机制的研究和认识。
在医学临床实践方面以Toll信号受体作靶向,可能研究和开发出防治某些重大疾病的新举措或高科技新药。比如,发现某种激活Toll样受体的无毒微生物成分介导上皮细胞高效表达抗菌肽,可能起到防治粘膜感染的作用;合成可激活Tlr-4信号受体的Spaetzle类似分子,或发现激活Tlr-4样受体的无毒微生物成分,或构建自发激活型Tlr-4样受体基因转染巨噬细胞,可能诱导抗肿瘤免疫反应;抗Tlr特异抗体或构建Toll样受体缺失基因转染相关细胞,以阻滞相关Toll样受体介导的信号传导,可能产生抗全身炎症反应综合征和动脉粥样硬化的治疗效应,等等。
, 百拇医药
参 考 文 献
[1] Vogel G. Fly development genes lead to immune find[J]. Science, 1998, 281: 1942~1944.
[2] 王伯瑶,吴 琦, 黄 宁. 天然免疫系统非克隆识别机制的进展[J]. 中国科学基金, 1999, 13:208~211.
[3] Johnston DSt, Nusslein-Volhard C. The origin of pattern and polarity in the drosophila embryo[J]. Cell, 1992, 68:201~219.
[4] Anderson KV, Nusslein-Volhard C. Information for the dorsal-ventral pattern of the Droshophila embryo is stored as maternal mRNA[J]. Nature, 1984, 311: 223~227.
, http://www.100md.com
[5] Anderson KV, Jurgens G, Nusslein-Volhard C. Establishment of Dorsal-ventral polatrity in the Drosophila embryo: genetic studies on the role of the Toll gene product[J]. Cell, 1985, 42: 779.
[6] Morisato D. Signaling pathways that establish the drosal-ventral pattern of the Drosophila embryo[J]. Annu Rev Genetics, 1995, 29: 371~399.
[7] Boman G. Peptide antibiotics and their role in innate immunity[J]. Annu Rev Immunol, 1995, 13:61~92.
, http://www.100md.com
[8] Lemaitre B, Nicolas E, Michaut L, et al. The Dorsoventral regulatory gene cassette spatzle/Toll/cactus controls the potent antifungal response in Drosophila adults[J]. Cell, 1996, 86:973~983.
[9] Tickle C. Worlds in common through NF-kappa B[J]. Nature, 1998, 392(6676): 547~549.
[10] Rock FL, Hardiman G, Timans JC, et al. A family of human receptors structurally related to Drosophila Toll[J]. Proc Natl Acad Sci USA, 1998, 95:588~593.
, http://www.100md.com
[11] Heguy A, Baldari CT, Macchiia G, et al. Amino acids conserved in interleukin-1 receptors(IL-1Rs) and the Drosophila Toll protein are essential for IL-1R signal transduction[J]. J Biol Chem, 1992, 267:2605~2609.
[12] Medzhitov R, Preston-Hur, Burt P & Janeway Jr CA. A human homolougues of the Drosophla Toll protein signals activation of adaptive immunity[J]. Nature, 1997, 388: 394~397.
[13] Yang R-B, Mark MR, Gray A. Toll-like receptor-2 mediates lipopolysaccharide-induced cellular signalling[J]. Nature, 1998, 395:284~287.
, 百拇医药
[14] Kirschning CJ, Wesche H, Ayres TM, et al. Human Toll-like receptor 2 confers responsiveness to bacterial lipopolysaccharide[J]. J Exp Med, 1998, 188: 2091~2097.
[15] Brightbill HD, Libraty DH, Krutzik SR, et al. Host defense mechanisms triggered by microbial lipoproteins through Toll-like receptors[J]. Science, 1999, 285: 732~736.
[16] Aliprantis AO, Yang R-B, Mark MR, et al. Cell activation and apoptosis by bacterial lipoproteins through Toll-like receptor-2[J]. Science, 1999, 285:736~739.
, 百拇医药
[17] Hoffman JA, Kafatos FC, Janeway Jr CA, et al. Phylogenetic perspectives in innate immunity[J]. Science, 1999, 284:1313~1318.
[18] Muzio M, Natoli G, Saccani S, et al. The human Toll signaling pathway: Divergence of nuclear factor κB and JNK/SAPK activation upstream of tumor necrosis factor receptor-associated factor 6(TRAF6)[J]. J Exp Med, 1998, 187: 2097~2101.
[19] Qureshi ST, Lariviere L, Leveque G, et al. Endotoxin tolerant mice have mutations in Toll-like receptor 4 (Tlr4)[J]. J Exp Med, 1999, 189:615~625.
, 百拇医药
[20] Hoshino K, Takeuchi O, Kawai T, et al. Cutting edge:Toll-like receptor 4 (Tlr 4)-deficient mice are hyporesponsive to lipopolysaccharide: evidence for Tlr 4 as the Lps gene product[J]. J Immunol, 1999, 162:3749~3752.
[21] Poterak A, He X, Smirnova I, et al. Defective LPS signaling in C3H/HeJ and C57BL/10ScCr mice:mutations in Tlr4 gene[J]. Science, 1998, 282: 2085~2088.
[22] Liao F, Andalibi A, deBeer FC, et al. Genetic control of inflammatory gene induction and NF-κB-like transcription factor activation in response to an atherogenic diet in mice[J]. J Clin Invest, 1993, 91:2572.
[收稿日期] 1999-09-28 [修回日期] 1999-11-04
, 百拇医药
单位:(华西医科大学感染免疫研究室 病理生理教研室, 四川 成都 610041)
关键词:炎症;信号传递;基因
中国病理生理杂志000625 [中图分类号] R364.5 [文献标识码] A
[文章编号] 1000-4718(2000)06-0567-06
Toll receptor signaling pathway in inflammation
WANG Bo-yao, HUANG Ning, WU Qi
(Research Unit of Infection & Immunity, Department of Pathophysiology, West China University of Medical Sciences, Chengdu 610041, China)
, 百拇医药
【A Review】 Toll signaling pathway may play a curcial role in induction of inflammation-associated gene activation. Originally, the Toll/spaetzle/Cactus-Dorsal signaling pathway is established in the Drosophila embryonic development. Recently, the Toll signaling pathway in adult Drosophila has been established in the induction of antimicrobial peptide expression. Five human Toll-like receptor genes (hTlr l-5) and one mouse Toll-like receptor gene (mTlr-4) have been isolated. Toll and Toll-like receptor genes encoded molecules are transmembrane proteins with an extracellular leucine repeat domain and a cytoplasmic domain homologous to IL-1 receptors. The intracellular signaling cascade involves Tube, Pelle, and Cactus-Dorsal complex in Drosophila, and MyD88, IRAK, TRAF 6, NIK, αβ-I κB kinase, and I κB -NFκB complex in mammals. Dorsal and NFκB are transcription factors, while Cactus and IκB are their inhibitors. When the inhibitors phosphorylated, the nuclear factors are released and move into nucleus, leading to immune gene activation. It has been shown from in vitro system that Tlr-4 mediated LPS signaling in human monocytes for expression of IL-1, IL-6, IL-8, and costimulator B7-1 which provides second signal for T cell response. Tlr-2 can also mediate LPS signaling in human monocytes, leading to the production of proinflammatory mediators. Microbial lipoproteins are potent stimulators of IL-12 production through Tlr-2 signaling by human macrophages, and can stimulate Tlr 2-dependent transcription of inducible nitric oxide synthase and the production of nitric oxide, a powerful microbicidal pathway. Findings of a point mutation of Tlr-4 in LPS tolerant C3H/HeJ mouse strain and a deletion of Tlr-4 in LPS resistant C57BL/10ScCr mice provide an in vivo evidence strongly supporting the crucial role of Tlrs in LPS mediated inflammation. It is proposed that targeting Tlrs would develop new remedies for treatment of inflammatory disorders and for immunotherapy of mucosal infections and cancer, etc.
, 百拇医药
[MeSH] Inflammation; Signal transduction; Genes
炎症反应是机体天然抵抗微生物侵袭的重要武器[1],因此从免疫学角度讲又称之为天然免疫。补体系统、凝血系统、急性期反应蛋白、吞噬细胞、内皮细胞和上皮细胞等构成这个反应系统的主要元件。天然免疫在发生学上是有机体最古老的抗感染防御机制,它使用胚胎细胞编码的形态识别受体(pattern recognition receptors, PRRs) 识别微生物的保守的分子成分,如LPS、胞壁脂蛋白、甘露糖和RNA病毒双链RNA等,产生抗微生物及其毒性因子的免疫反应[2]。这种分子形态识别受体从分布上可分为细胞性受体和体液性受体;从功能上可分为循环血中的体液性受体和在细胞膜上表达的内吞受体以及信号受体等。体液性受体如甘露糖结合蛋白、C反应蛋白、血清淀粉样蛋白等介导补体系统的激活和调理吞噬作用,而脂多糖结合蛋白(LBP)识别LPS,起浓集血中LPS并传递给CD14的作用,它们均属于由肝细胞合成的急性期反应蛋白;内吞受体如巨噬细胞甘露糖受体、清道夫受体和补体受体等介导吞噬作用;信号受体如CD14和Toll在细胞介导的炎症反应中起信号传导作用。天然免疫细胞识别病原体相关分子形态(pathogen associated molecular patterns, PAMs)或蛋白水解信号,一方面诱导抗菌肽的合成,直接杀伤侵入的病原微生物;另一方面诱导内源性信号的表达,如炎症预激细胞因子(TNF、IL-1等)、趋化细胞因子(IL-8、MCP、MIP等)、粘附分子、共刺激因子(B7、IL-12等)以及一氧化氮合酶(产生一氧化氮)等,以便更有效的动员机体的防御力量来抵抗病原微生物的侵入。人们已熟知髓性白细胞膜CD14介导LPS信号胞内传导而使NF-κB激活,以诱导上述内源性信号分子基因的表达。然而,CD14分子缺乏胞浆区段,不能直接向细胞内传导LPS信号,尚需其它分子起信号传导作用。新近发现的免疫调节基因产物Toll蛋白可能是其信号向胞内传导的门户蛋白(gateway),本文概要综述其研究进展。
, 百拇医药
一、从果蝇发育调节的研究发现免疫信号受体基因Toll
在果蝇胚胎的发育成形过程中最重要阶段是其受精卵向前后轴和背腹轴分化[3]。用遗传分析法在80年代发现了母体基因座Toll调控果蝇胚基细胞向背腹轴方向分化发育,形成胚胎的外胚层和中胚层[4,5]。以后用遗传学、分子生物学和生物化学方法确立了Toll基因产物Toll蛋白是分布在胚胎细胞上的信号受体,并确立了信号传导通路Spaetzle/Toll/Cactus-Dorsal[5]。Spaetzle基因编码47~60 kD蛋白质,分泌到卵黄周围间隙(卵黄膜和胚胎浆膜之间充满液体的空间),在4个丝氨酸蛋白酶家族成员级联反应作用下,最终使其水解产生23 kD的Spaetzle活性蛋白,作为Toll受体的配体而激活Toll蛋白,导致细胞内信号传导级联反应,即激活的Toll蛋白通过辅助因子Tube而使丝氨酸/苏氨酸蛋白激酶Pelle激活,Cactus的磷酸化和降解使转录因子Dorsal从无活性的胞浆蛋白复合物Cactus-Dorsal中释放出来,并移位到核内,激活相应的形态发生基因,导致果蝇胚胎细胞向背腹轴方向分化[6]。
, 百拇医药
在80年代初对果蝇抗感染机制的研究发现了抗菌肽,并被证明是果蝇抗感染的主要介质[7]。抗菌肽并不在果蝇体内固有表达,仅在感染性损伤或注射微生物成分和LPS时才在果蝇肥体细胞(在功能上相当于哺乳动物的肝细胞)和/或上皮细胞被诱导表达。现已知被诱导产生的抗菌肽分子至少有7种,即cecropins\, diptericin\, drosocin\, attacin\, 防御素和metchnikowin, 它们主要起抗细菌作用;而drosomycin具强力抗霉菌活性。所有这些抗菌肽基因的上游序列均含转录因子Dorsal结合位点,新近的实验研究已显示果蝇胚胎背腹轴成形级联反应调节基因Spaetzle/Toll/Cactus调控成年果蝇抗菌肽的表达[8]。Dorsal属于Rel蛋白家族成员,该蛋白家族是可迅速被诱导激活的基因的转录因子,在果蝇包括Dorsal和Dif蛋白;在哺乳动物是免疫和炎症反应效应基因转录激活因子NF-κB[9]。通过Database检索和分子同源性比较,发现IL-1诱导哺乳动物炎症反应中NF-κB激活的级联反应与果蝇Spaetzle/Toll/Cactus-Dorsal通路相似, 从而使学者们对Toll受体信号通路在哺乳动物天然免疫和炎症反应中的作用产生极大兴趣。现已克隆出5个人Toll样受体基因,称为Tlrs1-5(Toll-like receptors-1-5)[10],Tlrs又称hTolls。并相继在多个国际著名实验室和医药公司对其结构与功能进行热火朝天的研究[1]。
, 百拇医药
二、Toll蛋白的结构与功能的研究进展
Fig 1 Schematic structure of Toll and Toll-like receptors (from Rock FL, et al.Proc Natl Acad Sci USA. 1998, 95:588 with some modification)
图1 Toll及Toll样受体的基本结构
如图1所示,Toll属于I型跨膜受体蛋白,其细胞外区段(extracellular domain)由富含亮氨酸的重复序列和半胱氨酸结(半胱氨酸残基形成链内二硫键袢)所组成,果蝇Toll含2个半胱氨酸结,使其细胞外区段被第二个半胱氨酸结分隔成2个节段,而人Tlrs的细胞外区段仅含1个半胱氨酸结,紧邻细胞外膜;其细胞内胞浆区段(cytoplasmic domain)与哺乳动物IL-1受体的胞浆区段具同源性,果蝇Toll蛋白的胞浆区段的氨基酸序列与哺乳动物IL-1受体分子胞浆序列的同一性为26%,其相似性为43%[10,11]。然而,Toll蛋白的细胞外区段显然与IL-1受体相异,后者在结构上属Ig超家族。细胞膜CD14是一富含亮氨酸重复序列的糖基磷脂酰肌醇锚定蛋白,虽然与Toll蛋白细胞外区段在结构上具某种相似性,但缺乏胞浆区段,不能向细胞内传递信息。因此,Toll蛋白应为一类新的信号受体蛋白。
, 百拇医药
现克隆出的人Tlrs全长cDNAs分别为:Tlr-1, 2 366 bp,其开放读框编码786个氨基酸肽链;Tlr-2, 2 600 bp,其开放读框编码784个氨基酸肽链;Tlr-3, 3 029 bp, 其开放读框编码904个氨基酸肽链;Tlr-4, 3 811 bp,其开放读框编码879个氨基酸肽链;Tlr-5, 1 275 bp,其开放读框编码370个氨基酸肽链。以Tlrs cDNAs作探针进行荧光素标记原位杂交(FISH)染色体定位分析显示Tlr1-3基因定位于4号染色体上,Tlr4基因定位于9号染色体上,Tlr5基因定位于1号染色体上[10]。
Fig 2 Construction of an autologously activated Tlr4 chimaric gene (from Medzhitov R, et al. Nature, 1997, 388:394. AFR and CFR represent the amino-terminal flanking and carboxy-terminal cysteine-rich regions. LRR, leucine repeat region. TM, transmembrane region)
, 百拇医药
图2 自发激活型Tlr4嵌合基因的构建
关于Tlrs各成员的信号传递功能,现有报道显示Tlr-4跨膜段和胞浆区段与小鼠CD4胞外区段的编码基因片段构建的自身激活型嵌合基因,转染人髓系单核细胞系THP-1,无需配体即能激活NF-κB和表达炎症细胞因子IL-1、IL-6、IL-8及其共刺激分子B7-1(该分子为T细胞激活所必需的第二信号)[12]。该重组突变基因的构建见示意图2。
Tlr-2编码基因转染不表达Tlrs和CD14的人胚肾细胞系293,能使其接受LPS-LBP复合物刺激信号,诱导NF-κB激活和内皮选择素(E-selectin)基因的转录,而暂时共转染膜CD14编码基因的293细胞提高了对LPS反应敏感性和强度[13,14]。如图3所示,Tlr-2跨膜段和胞浆区段编码基因与CD4细胞外区段编码基因构建的CD4-Tlr-2嵌合基因,以及删除Tlr-2胞浆区段13位或141位氨基酸以后的编码序列而构建的缺失突变基因Tlr2-1和Tlr2-2,转染293细胞,对LPS反应性均丧失,前者表明Tlr-2细胞外区段为其识别LPS所必需的元件,后者提示其羧基端氨基酸序列为细胞内信号传导所必需,与IL-1受体膜内区段信号传导所必需的氨基酸同一性很高,见图3。
, 百拇医药
Fig 3 Construction of deletion mutants of Tlr-2 gene (from Yang R-B, et al. Nature 1998, 395:284.ECD, extracellular domain. ICD, intracellular domain. TM, transmembrane region)
图3 Tlr-2缺失突变基因的构建
由于人胚肾293细胞系既不表达CD14和Tlrs也不表达炎症细胞因子,已作为一个体外模式系统用于人Tlrs基因的功能研究。最新研究报道[15]显示细菌胞壁脂蛋白,如结核杆菌19 kD脂蛋白、burgdorferi包柔氏螺旋体脂蛋白(OspA)和梅毒螺旋体47 kD脂蛋白等,能刺激转染了Tlr-2编码基因的293细胞系的IL-12基因的转录激活,共转染CD14编码基因加强这种转录激活作用;Tlr-2胞浆区段缺失突变基因转染RAW264.7巨噬细胞系,抑制细菌脂蛋白介导的IL-12和一氧化氮合酶基因的转录激活。亦有报道细菌脂蛋白能介导转染了Tlr-2编码基因的293细胞的凋亡,提示Tlrs可能在炎症反应的负相调节机制中发挥作用[16]。
, 百拇医药
三、感染性炎症反应的Toll信号转导通路[12, 17, 18]
Fig 4 Toll and Toll-like receptor-mediated intracellular signaling (From Hoffman JA, et al. Science, 1999, 284:1313 with a little modification)
图4 Toll及Toll样受体介导的细胞内信号传递
如图4所示,左边代表Toll信号通路诱导成年果蝇抗霉菌基因表达抗霉菌多肽drosomycin,右边代表LPS-LBP-CD14复合物与人Tlr-4结合而激活共刺激因子B7基因的胞内信号传导通路。当成年果蝇受到霉菌感染时,通过其尚未明了的体液性形态识别受体与病原体相关形态分子结合而触发淋巴液中一系列酶解反应,产生激活型Spaetzle而激活Toll蛋白,引起胞内信号传导级联反应,这个反应涉及Tube蛋白、丝氨酸/苏氨酸激酶Pelle,最终使Cactus磷酸化而降解,释放转录因子Dorsal,激活的Dorsal向细胞核移位,与抗霉菌基因增强子序列中的顺式作用元件结合而使其激活,合成抗霉菌多肽。显然该通路与调控果蝇胚胎早期发育背腹轴细胞分化的Toll信号传导通路一致。LPS-LBP-CD14 复合物与人单核吞噬细胞Tlr-4结合触发胞内级联信号传导反应,首先Tlr-4胞浆区段与一个叫做衔接子(adapter)的蛋白MyD88相互作用,依次激活IL-1受体相关激酶(IRAK)、肿瘤坏死因子受体相关因子-6(TRAF6)、NF-κΒ诱导激酶(NIK)和αβ-IκB激酶,最终使IκB磷酸化而降解,激活的NF-κB移位入核内,启动免疫反应基因转录激活。在哺乳动物Toll介导的胞内信号传导所涉及的重要分子无论在结构上或在功能上均与成年果蝇Toll介导的免疫信号胞内传导通路(如NF-κB与Dorsal; IκB与Cactus; IRAK与Pelle等)具相似性,说明Toll介导的免疫和炎症信号传导通路起源古老和保守。
, http://www.100md.com
四、Toll基因突变与内毒素耐受性[19~21]
依据人Tlr-4基因序列设计引物,应用逆转录-聚合酶链反应(RT-PCR)技术已分离出小鼠Tlr-4 cDNA,其编码的氨基酸序列与人Tlr-4具高度同源性(同一性为67%, 相似性为79%),而胞浆区段的同源性比胞外区段高(前者为89%,后者为68%)。
人们已熟知细菌LPS诱发感染宿主强烈的全身炎症反应。在小鼠,这一反应的遗传调节定位于4号染色体Lps基因座,存在2个等位基因,即LPS低反应基因(Lpsd)和LPS反应基因(Lpsn)。C3H/HeJ和C57BL/10ScCr是Lpsd-d纯合子小鼠系,它们对LPS的攻击不发生全身炎症反应,即对LPS具耐受性,然而其分子遗传机制尚未明了。最新研究报道用遗传学和限制性内切酶作图技术确定Lps基因座图距为0.9厘摩(cM),核苷酸序列跨距为1.7 Mb,包含3个转录单位,其中之一为Tlr-4基因。C3H/HeJ小鼠系Tlr-4基因编码区发生了点突变,即712位上编码脯氨酸的密码子由编码组氨酸的密码子替换了,其结果是合成的蛋白分子712位上(位于Tlr-4肽链的胞浆区段)的脯氨酸变成了碱性组氨酸,推测这一氨基酸的突变可能使Tlr-4胞浆区段构型发生改变而不能与下游分子发生相互作用,致使LPS通过Tlr-4蛋白介导的胞内信号传导被中断。而在C57BL/10ScCr小鼠系Lps基因座中却表现Tlr-4基因的缺失。这些研究结果提示Tlr-4结构与功能的变化与机体对内毒素的耐受性密切相关,另一方面从整体水平上提供了证据表明Tlrs信号受体在介导炎症刺激信号的胞内传导中起重要作用。
, 百拇医药
五、小结与展望
在果蝇胚胎发育研究中发现其背腹成形受Toll基因的调控。新近发现该基因亦是病原微生物诱导成年果蝇产生抗菌肽的信号传导通路的门户蛋白基因,从人亦克隆出5个Toll样蛋白家族成员,从小鼠克隆出一个与人Tlr-4具高度同源性的基因。体外和体内实验研究均显示Tlr-4介导内毒素信号胞内传导,激活免疫和炎症相关基因,产生相应的细胞因子或其它介质。在体外实验系统,亦显示Tlr-2可介导内毒素和细菌脂蛋白信号的胞内传导。这些实验结果提供了一个有力证据表明炎症反应与生物的发育、进化发展密切相关,即发育调控基因的趋异(divergent evolution)进化分化出免疫调节基因,以抵抗病原微生物的侵袭,保证复细胞生物的发育和进化。天然免疫系统起源古老,其调控分子相当保守。因此,研究Toll及其相关基因结构与功能可能是揭示哺乳动物和人炎症反应的本质特征的一个突破口。
无论是感染性炎症反应或非感染性慢性炎症所导致的组织器官损伤和疾病状态,如全身炎症反应综合征所致的休克及多器官功能衰竭、进行性毁损性青少年牙周炎、动脉粥样硬化等,均只在一定的易感人群中发生,提示可能有其遗传背景,C3H/HeJ小鼠系对高脂食物诱发动脉粥样硬化具耐受性[22]已提供证据支持这种假设。在对内毒素耐受小鼠系C3H/HeJ和C57BL/10ScCr上的分子遗传研究显示Tlr-4基因的突变是其内毒素耐受的原因,这为揭示上述炎症疾病的分子遗传基础的研究提供了一条令人鼓舞的途径。
, http://www.100md.com
从信号受体Toll蛋白基因的研究显示,炎症反应是低等动物发育过程中分化出的抗微生物感染的天然防御机制。在低等的无脊椎动物如昆虫,其生物学效应主要是产生抗菌肽,以行使抗感染作用;随进化发展,到了哺乳动物和人,炎症反应分化发展得相当复杂了。有体液反应系统和细胞介导系统;在细胞介导系统中,有被诱导产生的各种炎症细胞因子、其它小分子炎症介质以及抗菌肽;其反应或强或弱;既可产生保护效应又可发生损伤后果。深入研究古老的Toll及其相关基因的结构和功能可能对这一复杂反应理出一定头绪。比如,抗菌肽的产生和对细胞介导反应的正相与负相调节起作用的各种内源性信号的表达可能由不同的Toll样信号受体介导; 在体液反应系统的级联反应酶解产物中可能存在Spaetzle相关分子,这方面的研究有可能把体液反应系统与细胞介导系统联系起来,且可推进和加深对非感染性炎症疾病如自身免疫疾病和动脉粥样硬化发病机制的研究和认识。
在医学临床实践方面以Toll信号受体作靶向,可能研究和开发出防治某些重大疾病的新举措或高科技新药。比如,发现某种激活Toll样受体的无毒微生物成分介导上皮细胞高效表达抗菌肽,可能起到防治粘膜感染的作用;合成可激活Tlr-4信号受体的Spaetzle类似分子,或发现激活Tlr-4样受体的无毒微生物成分,或构建自发激活型Tlr-4样受体基因转染巨噬细胞,可能诱导抗肿瘤免疫反应;抗Tlr特异抗体或构建Toll样受体缺失基因转染相关细胞,以阻滞相关Toll样受体介导的信号传导,可能产生抗全身炎症反应综合征和动脉粥样硬化的治疗效应,等等。
, 百拇医药
参 考 文 献
[1] Vogel G. Fly development genes lead to immune find[J]. Science, 1998, 281: 1942~1944.
[2] 王伯瑶,吴 琦, 黄 宁. 天然免疫系统非克隆识别机制的进展[J]. 中国科学基金, 1999, 13:208~211.
[3] Johnston DSt, Nusslein-Volhard C. The origin of pattern and polarity in the drosophila embryo[J]. Cell, 1992, 68:201~219.
[4] Anderson KV, Nusslein-Volhard C. Information for the dorsal-ventral pattern of the Droshophila embryo is stored as maternal mRNA[J]. Nature, 1984, 311: 223~227.
, http://www.100md.com
[5] Anderson KV, Jurgens G, Nusslein-Volhard C. Establishment of Dorsal-ventral polatrity in the Drosophila embryo: genetic studies on the role of the Toll gene product[J]. Cell, 1985, 42: 779.
[6] Morisato D. Signaling pathways that establish the drosal-ventral pattern of the Drosophila embryo[J]. Annu Rev Genetics, 1995, 29: 371~399.
[7] Boman G. Peptide antibiotics and their role in innate immunity[J]. Annu Rev Immunol, 1995, 13:61~92.
, http://www.100md.com
[8] Lemaitre B, Nicolas E, Michaut L, et al. The Dorsoventral regulatory gene cassette spatzle/Toll/cactus controls the potent antifungal response in Drosophila adults[J]. Cell, 1996, 86:973~983.
[9] Tickle C. Worlds in common through NF-kappa B[J]. Nature, 1998, 392(6676): 547~549.
[10] Rock FL, Hardiman G, Timans JC, et al. A family of human receptors structurally related to Drosophila Toll[J]. Proc Natl Acad Sci USA, 1998, 95:588~593.
, http://www.100md.com
[11] Heguy A, Baldari CT, Macchiia G, et al. Amino acids conserved in interleukin-1 receptors(IL-1Rs) and the Drosophila Toll protein are essential for IL-1R signal transduction[J]. J Biol Chem, 1992, 267:2605~2609.
[12] Medzhitov R, Preston-Hur, Burt P & Janeway Jr CA. A human homolougues of the Drosophla Toll protein signals activation of adaptive immunity[J]. Nature, 1997, 388: 394~397.
[13] Yang R-B, Mark MR, Gray A. Toll-like receptor-2 mediates lipopolysaccharide-induced cellular signalling[J]. Nature, 1998, 395:284~287.
, 百拇医药
[14] Kirschning CJ, Wesche H, Ayres TM, et al. Human Toll-like receptor 2 confers responsiveness to bacterial lipopolysaccharide[J]. J Exp Med, 1998, 188: 2091~2097.
[15] Brightbill HD, Libraty DH, Krutzik SR, et al. Host defense mechanisms triggered by microbial lipoproteins through Toll-like receptors[J]. Science, 1999, 285: 732~736.
[16] Aliprantis AO, Yang R-B, Mark MR, et al. Cell activation and apoptosis by bacterial lipoproteins through Toll-like receptor-2[J]. Science, 1999, 285:736~739.
, 百拇医药
[17] Hoffman JA, Kafatos FC, Janeway Jr CA, et al. Phylogenetic perspectives in innate immunity[J]. Science, 1999, 284:1313~1318.
[18] Muzio M, Natoli G, Saccani S, et al. The human Toll signaling pathway: Divergence of nuclear factor κB and JNK/SAPK activation upstream of tumor necrosis factor receptor-associated factor 6(TRAF6)[J]. J Exp Med, 1998, 187: 2097~2101.
[19] Qureshi ST, Lariviere L, Leveque G, et al. Endotoxin tolerant mice have mutations in Toll-like receptor 4 (Tlr4)[J]. J Exp Med, 1999, 189:615~625.
, 百拇医药
[20] Hoshino K, Takeuchi O, Kawai T, et al. Cutting edge:Toll-like receptor 4 (Tlr 4)-deficient mice are hyporesponsive to lipopolysaccharide: evidence for Tlr 4 as the Lps gene product[J]. J Immunol, 1999, 162:3749~3752.
[21] Poterak A, He X, Smirnova I, et al. Defective LPS signaling in C3H/HeJ and C57BL/10ScCr mice:mutations in Tlr4 gene[J]. Science, 1998, 282: 2085~2088.
[22] Liao F, Andalibi A, deBeer FC, et al. Genetic control of inflammatory gene induction and NF-κB-like transcription factor activation in response to an atherogenic diet in mice[J]. J Clin Invest, 1993, 91:2572.
[收稿日期] 1999-09-28 [修回日期] 1999-11-04
, 百拇医药