ЮЂбЛЗЯЕЭГОжВППижЦЙ§ГЬЕФЩњЮяСІбЇФЃаЭ
зїепЃКЙљжйШ§
ЕЅЮЛЃКЙљжйШ§(жаЙњПЦбЇдК ГЩЖММЦЫуЛњгІгУбаОПЫљздЖЏЭЦРэЪЕбщЪвЃЌГЩЖМ 610041)
ЙиМќДЪЃКЮЂбЛЗЃЛОжВППижЦЃЛЩњЮяСІбЇФЃаЭ
ЩњЮявНбЇЙЄГЬбЇдгжО000204 еЊвЊ ЗжЮіСЫМфжЪФкбЊЙмДњаЛЮяжЪЕФВњЩњКЭдЫЖЏЁЂУЋЯИбЊЙмЧАРЈдММЁЕФПЊБеЙ§ГЬЁЂЮЂЖЏТіЦНОљАыОЖЕФгАЯьвђЫиМАЮЂЖЏТіздТЩдЫЖЏеёЗљЕФБфЛЏЧщПіЃЌНЈСЂСЫЮЂбЛЗЯЕЭГОжВППижЦЙ§ГЬЕФЩњЮяСІбЇФЃаЭЃЌгажњгкЖдИУЙ§ГЬНјааЖЈСПЗжЮіЁЃ
A Biomechanical Model of Local Control of Microcirculation
Guo Zhongsan
(Chengdu Institute of Computer Application, Academia Sinica, Chengdu 610041)
, http://www.100md.com
Abstract In this paper we analyze the generation and motion of vasoactive metabolites in interstitium, the open and closure of precapillary sphincters, the factors affecting mean arteriolar diameter, and the change of amplitude of arteriolar vasomotion. A biomechanical model of local control of microcirculation has been developed. It is useful in quantitative analysis of microcirculatory local control.
Key words Microcirculation Local control Biomechanical model
, АйФДвНвЉ
1 в§ бд
ЮЂбЊЙмДВЪЧзщжЏЙрСїЁЂбЊвК-зщжЏСїЬхНЛЛЛКЭзщжЏбЊШнСПЕФПижЦГЁЫљЁЃжївЊЕФПижЦепЪЧЮЂЖЏТіКЭУЋЯИбЊЙмЧАРЈдММЁЃЌжївЊПижЦЛњжЦгаОжВППижЦ(Local control,ЛђIntrinsic control)КЭШЋЩэПижЦ(Remote control,ЛђExtrinsic control)ЃЌжївЊПижЦФПБъЪЧдкИїжжЛЗОГЬѕМўИФБф(зюГЃМћЕФЪЧбЊбЙКЭбѕХЈЖШИФЖШ)ЪББЃГжФкЛЗОГЕФЮШЖЈадЁЃ
ШЋЩэПижЦжївЊжИЩёОПижЦгыЬхвКПижЦЁЃЖўепЙиЯЕЪЎЗжУмЧаЃЌвђЮЊЩёОПижЦвВЪЧЭЈЙ§ЬхвКвђзгЪЕЯжЕФ[1]ЃЌЬхвКвђзгЕФДѓВПЗжЪЧЩёОЕнжЪЃЌШчШЅМзЩіЩЯЯйЫиЁЂввѕЃЕЈМюЕШЃЌгаЕФдђЪЧЯИАћЕФЪЭЗХВњЮяЃЌШчЯйпА(adenosine)ЁЃ
ОжВППижЦАќРЈМЁдадПижЦгыДњаЛадПижЦЁЃМЁдадПижЦЪЧжИЮЂЖЏТідкФкЭтбЙВюдіДѓЪБжїЖЏЪеЫѕвдПижЦбЊСїСПЕФЯжЯѓЁЃДњаЛадПижЦЪЧЭЈЙ§ЯИАћДњаЛВњЮяНјааЕФПижЦЁЃгабаОПШЯЮЊ[1]ЃЌЮЂбЛЗЕФОжВПЕїНкжаЃЌ75%ЪЧДњаЛадЕїНкЁЃ
, http://www.100md.com
ЮЂбЛЗЯЕЭГПижЦЙ§ГЬЕФЖЈСПбаОПЙЄзїЗЧГЃЩйЁЃзїепЗжЮібаОПСЫгаЙиЕФЩњРэЙ§ГЬЃЌНЈСЂСЫЮЂбЛЗЯЕЭГОжВППижЦЙ§ГЬЕФЩњЮяСІбЇФЃаЭЃЌгаРћгкЖдетвЛЙ§ГЬНјааЖЈСПЗжЮіЁЃ
2 МфжЪЯйпАдДЧПЖШМАЯйпАХЈЖШЗжВМЙЋЪН
ЯИАћ(МйЩшОљдШЗжВМдкМфжЪПеМфФк)ЪЭЗХЛюбЊЙмДњаЛЮяжЪ(дкаФМЁжаЃЌзюживЊЕФЪЧЯйпАЃЛЮЊа№ЪіЗНБуЃЌБОЮФжаЮвУЧгУЯйпАДњБэЫљгаПЩФмЕФЛюбЊЙмДњаЛЮяжЪ)ШЁОігкЯпСЃЬхЕФбѕРћгУТЪЁЃЯпСЃЬхФкЕФЯИАћЩЋЫи(Cytochrom)aa3вдЛЙдгыбѕЛЏСНжжЗНЪНДцдкВЂПЩЛЅЯрзЊЛЏЁЃбѕНјШыЯИАћКѓЃЌМДзїЮЊЕчзгЕФЪмЬхгызїЮЊЕчзгЙЉЬхЕФЛЙдaa3ЯрЛЅзїгУЃЌЖјЯпСЃЬхЕФбѕРћгУТЪUO2MMОЭгЩЯИАћФкЕчзгЪмЬхЕФХЈЖШCO2cellcellгыЕчзгЙЉЬхЕФХЈЖШ[aa3]RЙВЭЌОіЖЈ[2]ЃК (1)
(1)
, АйФДвНвЉ
дкОВЯЂзДЬЌЯТЃЌ[aa3]RжЛеМЯИАћЩЋЫизмХЈЖШ[aa3]TЕФ1/10ЛђИќЩйЁЃЕБЯИАћбѕХЈЖШНЕЕЭЪБЃЌбѕЛЏзїгУМѕШѕЃЌaa3ЕФбѕЛЏ-ЛЙдЖЏЬЌЦНКтБЛЦЦЛЕЃЌВПЗжбѕЛЏЕФaa3БЛЛЙдЃЌ[aa3]RдіМгЃЌДгЖјВЙГЅСЫCO2cellcellНЕЕЭЕФгАЯьЃЌЪЙUO2MMЮЌГжВЛБфЛђЛљБОВЛБфЁЃетжжВЙГЅзїгУНјааЕНaa3ШЋВПЛЙдЪБЃЌЯИАћбѕХЈЖШОЭЕНСЫвЛИіСйНчжЕCO2ccccЃЌНјвЛВННЕЕЭОЭЛсЪЙUO2MMЯджјМѕаЁЃЌЯИАћПЊЪМДѓСПЪЭЗХЯйпАЁЃОнДЫПЩЕУ[2]ЃК (2)
(2)
, АйФДвНвЉ
ЦфжаЃКPO2ЊЊcellЮЊЯИАћбѕЗжбЙЃЛUO2ЊЊmaxЮЊЯпСЃЬхзюДѓбѕРћгУТЪЃЌГЃСПЃЛKMЮЊЯИАћбѕЗжбЙВЮПМжЕЃЌБэЪОЯпСЃЬхбѕРћгУТЪЮЊзюДѓбѕРћгУТЪЕФ50%ЪБЕФЯИАћбѕЗжбЙЁЃдкДѓЖрЪ§зщжЏжаЃЌKM<0.1 mmHgЁЃЖјЭЈГЃЯИАћбѕЗжбЙБШKMДѓ1ЁЋ2ИіЪ§СПМЖ[37ЁцЪБдМЮЊ2ЁЋ5 mmHg(1mmHg=0.133kPa)]ЁЃвђДЫЃЌЯИАћбѕХЈЖШдкЯрЕБДѓЕФЗЖЮЇФкБфЛЏЪБЃЌUO2ЊЊMЕФБфЛЏЖМЗЧГЃаЁЁЃвЛАуЕиЫЕЃЌЯИАћбѕЗжбЙДѓгк1 mmHgЪБЃЌОљПЩНќЫЦШЯЮЊUO2ЊЊMЪЧвЛИіГЃСПЁЃдкБОЮФжаЃЌЮвУЧгУЯпСЃЬхЯрЖдбѕРћгУТЪ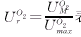
, http://www.100md.com
БэЪОЯИАћЪЧЗёШБбѕМАШБбѕГЬЖШЃЛР§ШчЃЌUO2rr<0.95ЮЊЧсЖШШБбѕЃЌUO2rr<0.90ЮЊжиЖШШБбѕЃЌЕШЕШЁЃ
ЯИАћбѕХЈЖШгЩЯТЪНИјГі[3]ЃК
ЦфжаЃКVcellЮЊЯИАћЬхЛ§ЃЌJO2cellcellЮЊНјШыЯИАћЕФбѕЭЈСПЃК
ЦфжаЃКCO2IIЮЊМфжЪбѕХЈЖШЁЃгЩгкЖдбѕЗжзгРЉЩЂЕФзшСІЃЌЯИАћФкЭтДцдкбѕХЈЖШЬнЖШЁЃЪЕбщБэУїЃЌЯИАћОпгаЕїНкетвЛбѕХЈЖШЕФФмСІЃЌдкМфжЪШБбѕЪБМѕаЁетвЛЬнЖШЃЌДгЖјВПЗжЕиЮЌГжЯИАће§ГЃЕФбѕКЯзїгУ[4]ЁЃетжжЙІФмадБфЛЏПЩФмЪЧвђЮЊaa3гыбѕЕФЧзКЯСІЫцЯИАћЕФФмСПзДПіЗЂЩњИФБфЫљв§Ц№ЕФ[5]ЁЃвђДЫЃЌЩЯЪНжаЕФЯЕЪ§Kc2гІИУЪЧ[aa3]RЕФКЏЪ§ЁЃЕЋдкБОЮФжаЃЌЮвУЧднШЁKc2ЮЊГЃСПЁЃ
, http://www.100md.com
ПЩвдПДГіЃЌЯпСЃЬхбѕРћгУТЪЁЂЯИАћбѕХЈЖШКЭМфжЪбѕХЈЖШЕФЙиЯЕЪЧКмИДдгЕФЁЃзїЮЊМђЛЏЃЌЮвУЧМйЩшЯИАћбѕХЈЖШЫцЪБМфБфЛЏКмТ§ЃЌНјШыЯИАћЕФбѕгыЯпСЃЬхЯћКФЕФбѕЫцЪББЃГжЦНКтЃЌДгЖјЕУЕНЃК
ЦфжаЃК ЮЊЖдгІгкKMЕФбѕХЈЖШЁЃ
ЮЊЖдгІгкKMЕФбѕХЈЖШЁЃ
ИљОнЩЯЪіЗжЮіЃЌВЂМйЩшЯИАћЪЭЗХЯйпАЕФЧПЖШгыЯпСЃЬхбѕРћгУТЪГЩЯпадЙиЯЕЃЌЮвУЧЕУЕНШчЯТЯйпАдДЧПЖШЙЋЪНЃК (3)
(3)
ЦфжаЃКSadЮЊМфжЪЯйпАдДЧПЖШЃЛKadЮЊЯйпАдДЧПЖШЯЕЪ§ЁЃ
ДгЯйпАдДВњЩњЕФЯйпАдкМфжЪФкЕФЪфдЫЙ§ГЬгыбѕЯрЫЦЃЌвВАќРЈРЉЩЂЁЂЖдСїКЭЛњаЕУжЩЂШ§ЗНУцЃЌЧвврПЩМйЩшМфжЪЖдЯйпАЮоХХГтзїгУЁЃДЫЭтЃЌбЊвКбЛЗКЭУЋЯИбЊЙмЭјЕФДцдкЖдЯйпАЕФЗжВМКЭдЫЖЏФмВњЩњКмДѓгАЯьЁЃЪЕбщБэУїЃЌдкЮЂОВТізЂШы0.1 ІЬMХЈЖШЕФЯйпАЃЌФмЯджјРЉеХЯрСкЮЂЖЏТіЕФЙмОЖ[6]ЃЌЫЕУїдкЮЂЖЏТі-ЮЂОВТіжЎМфДцдкЁАРЉЩЂНЛСЌЁБЃЌЪЙвбОЁАРыПЊЁБСЫЮЂбЛЗЕЅдЊЕФЯйпАШдФмВњЩњгАЯьЁЃдкУЋЯИбЊЙмФкзЂШыШЅМзЩіЩЯЯйЫиЃЌФмЪЙЙЉбЊЮЂЖЏТіЙмОЖМѕаЁЃЌУЋЯИбЊЙмСїЫйНЕЕЭ(жБжСНЕЮЊСу)ЃЛзЂШыввѕЃЕЈМяЛђЛКМЄыФЃЌПЩЪЙЮЂЖЏТіРЉеХВЂдіМгбЊСїЫйЖШ[7ЃЌ8]ЁЃзїепШЯЮЊЃЌетжжДгУЋЯИбЊЙмЕНЮЂЖЏТіЕФЭЫааадЕїНкЃЌМШВЛЪЧДгУЋЯИбЊЙмжБНгЯђЮЂЖЏТіРЉЩЂЕФНсЙћЃЌвВВЛЪЧЭЈЙ§ЮЂЖЏТі-ЮЂОВТіРЉЩЂНЛСЌЪЕЯжЕФЃЌЖјЪЧвђЮЊУЋЯИбЊЙмБОЩэОЭОпгаДЋЪфЛюбЊЙмЮяжЪЕФЙІФмЁЃДЫЭтЃЌУЋЯИбЊЙмФкЦЄЯИАћЛЙФмЮќЪеДѓСПЯйпА[9]ЁЃвђДЫЃЌЛюбЊЙмЮяжЪдкЮЂбЛЗЕЅдЊжаЕФдЫЖЏЗНЪНПЩФмЪЎЗжИДдгЃЌгаД§НјвЛВНбаОПЁЃзїЮЊМђЛЏЃЌзїепгУдДЕуЕќМгЗЈМЦЫуМфжЪИїДІЕФЯйпАХЈЖШЃЌiЕуЕФЯйпАдДSadiдкjЕуВњЩњЕФЯйпАХЈЖШCadI(ij)гЩЦНЗНЗДБШТЩИјГіЃК
, http://www.100md.com
ЦфжаЃКXijЮЊiЁЂjжЎМфЕФОрРыЁЃетбљЃЌФГвЛЪБПЬjЕуЕФЯйпАХЈЖШОЭгЩЯТЪНИјГіЃК (4)
(4)
ЦфжаЃКCadI(j)ЮЊМфжЪФкФГЕуjЕФЯйпАХЈЖШЃЛSadiЮЊФГЕуiЕФЯйпАдДЧПЖШЁЃЦфЛ§ЗжаЮЪНЮЊЃК
ЦфжаЃКVЮЊМфжЪЬхЛ§ЁЃ
3 УЋЯИбЊЙмЧАРЉдММЁЕФОжВППижЦФЃаЭ
НќДњЕФбаОПБэУї[10ЃЌ11]ЃЌУЋЯИбЊЙмЕФДЂБИ(Reserve)гыФММЏ(Recruitment)ЬсЙЉСЫвЛИіОЋЧЩЕФЕїНкЛњжЦЃЛзщжЏФкЙиБезХЕФУЋЯИбЊЙмашвЊЪБПЩвдПЊЗХЃЌПЊЗХзХЕФУЋЯИбЊЙмврПЩФмЙиБеЃЌЪЙбЊвКдкЮЂбЊЙмДВФкИќгааЇЕижиаТЗжВМЃЌДгЖјдкбЊСїСПВЛБфЕФЬѕМўЯТБуПЩТњзудіМгСЫЕФашвЊЁЃетжжЕїНкЪЧЮЂбЛЗЕЅдЊФкВПЕФЕїНкЃЌВЛгАЯьЦфгрЕЅдЊЕФЙрСїЃЌВЛдіМгаФдрБУбЊЙІФмЃЌдкБОЮФжаЮвУЧГЦжЎЮЊФкВПЕїНкЛђЕквЛЕїНкЁЃ
, http://www.100md.com
ЕБбЊвКЗжВМдкЫљгаПЩНЛЛЛУцЩЯШдВЛФмТњзузщжЏЕФашвЊЪБЃЌЮЂЖЏТіНЋЛсЯджјЕиРЉеХЃЌЪЙНјШыИУЮЂбЛЗЕЅдЊЕФбЊСїСПдіМгЃЌДгЖјТњзузщжЏЕФашвЊЁЃетжжЕїНкЗНЪНЛсИФБфЦфгрЮЂбЛЗЕЅдЊЕФбЊСїзДПіЃЌЛђдіМгаФдрИККЩЃЌвђДЫЮвУЧГЦжЎЮЊЭтВПЕїНкЛђЕкЖўЕїНкЁЃ
ЩшУЋЯИбЊЙмЧАРЈдММЁДІЕФЯйпАХЈЖШДѓгкЕквЛЕїНкСйНчжЕCmlЪБЃЌИУУЋЯИбЊЙмМДЭъШЋПЊЗХЃЌЗДжЎдђЭъШЋЙиБеЃЌБуЕУЕНШчЯТЕФУЋЯИбЊЙмЧАРЈдММЁДњаЛадЕїПиФЃаЭЃК (5)
(5)
ЦфжаЃКrЊcЮЊУЋЯИбЊЙмАыОЖЃЛrЊcoЮЊАыОЖГЃСПЃЛJ(x)ЮЊНздОКЏЪ§ЁЃ
4 ЮЂЖЏТіЦНОљАыОЖЕФОжВППижЦФЃаЭ
ЮЂЖЏТіМЁдадПижЦЕФЛњРэЛЙВЛКмЧхГўЃЌФПЧАЩаЮоЦеБщНгЪмЕФЩњЮяСІбЇФЃаЭЁЃЪЕМЪгІгУжаГЃВЩгУLaplaceЙЋЪНЃК
, http://www.100md.com
rp=ГЃЪ§
ЦфжаЃКrЮЊЮЂЖЏТіАыОЖЃЛpЮЊбЙСІЁЃетЪЧИљОнЮЂЖЏТіБкФкКЯСІВЛБфЕФМйЩшЖјЕУЕНЕФЁЃгЩгкЮЂЖЏТіЪєКёБкЙмЃЌЙЪЮвУЧМйЩшМЁдадЕїНкЕФФПЕФЪЧБЃГжЮЂЖЏТіБкФкЦНОљгІСІІвaВЛБфЃЌМДЃК
rp=Івah hЁЊБкКё
ЭЌЪБМйЩшЙмБкВЛПЩбЙЫѕЃЌгЩДЫЕУЕНЮЂЖЏТіЕФМЁдадЕїПиФЃаЭЃК (6)
(6)
ЦфжаЃКa0ЮЊЮЂЖЏТіШыПкАыОЖЦНОљжЕЃЛa00ЮЊе§ГЃЧщПіЯТЮЂЖЏТіШыПкАыОЖЦНОљжЕЃЛPa0ЮЊЮЂЖЏТіШыПкДІбЊбЙЃЛPЁфaoЮЊе§ГЃЧщПіЯТЮЂЖЏТіШыПкДІбЊбЙЃЛ ЃЛh0ЮЊЮЂЖЏТіШыПкДІЙмБкКёЖШЁЃгЩгкЪНжаАќКЌСЫАыОЖгыБкКёжЎБШЃЌЙЪБШLaplaceЙЋЪНИќФмЗДгГВЛЭЌЮЂЖЏТіЕФВюБ№ЁЃПЩвдПДГіЃЌІвaдНаЁЃЌЩЯЪНдННгНќгкLaplaceЙЋЪНЁЃ
ЃЛh0ЮЊЮЂЖЏТіШыПкДІЙмБкКёЖШЁЃгЩгкЪНжаАќКЌСЫАыОЖгыБкКёжЎБШЃЌЙЪБШLaplaceЙЋЪНИќФмЗДгГВЛЭЌЮЂЖЏТіЕФВюБ№ЁЃПЩвдПДГіЃЌІвaдНаЁЃЌЩЯЪНдННгНќгкLaplaceЙЋЪНЁЃ
, АйФДвНвЉ
ДгВЛЭЌЕФНЧЖШПЩвдИјГіВЛЭЌЕФФЃаЭЁЃР§ШчЃЌгаЕФзїеп[12]ЭЈЙ§в§ШыЁАВЮПМжБОЖЁБЕФИХФюРДНЈСЂЮЂЖЏТіМЁдадЕїПиФЃаЭЁЃжЕЕУжИГіЕФЪЧЃЌИУЮФзїепЕУЕНЕФrЫцPЕФБфЛЏЫйЖШКЭДЫДІЕФ(6)ЪНЖМБШLaplaceЙЋЪНЫљИјГіЕФЫйЖШТ§ЁЃ
ЮЂЖЏТіАыОЖЫцЮЂЖЏТіШыПкДІЕФЯйпАХЈЖШдіМѕЖјдіМѕЁЃИљОнЖдН№ЛЦВжЪѓЕФЪЕбщ[13]ЃЌЕБЯйпАХЈЖШЮЊ10-4MзѓгвЪБЃЌЮЂЖЏТіРЉеХДяЕНзюДѓжЕЁЃИїМЖЮЂЖЏТіЕФОВЯЂжБОЖдМЮЊзюДѓжБОЖЕФ70%ЁЃЕБзщжЏбѕХЈЖШГЌЙ§ашвЊЪБЃЌЮЂЖЏТіЙмОЖНЋМѕаЁЃЌДгЖјЬсЙЉвЛИіИКЗДРЁЕїНкЃЌЪЙзщжЏбѕЙЉгІгыЯћКФЧїгкЦНКтЁЃЕЋдкЖЈСПбаОПЩЯФПЧАДцдкНЯДѓВюБ№ЃКгаЕФШЯЮЊЖЏТібѕЗжбЙИпгке§ГЃжЕЪБЃЌЮЂЖЏТіжБОЖНігаКмаЁЕФМѕЩй[14]ЃЌЖјKunertЕШ[15]гУНЕЕЭбЊЧђбѕЧзКЭСІЕФЗНЗЈЕУЕНЕФНсЙћдђЪЧЕБзщжЏбѕЗжбЙдіМгАйЗжжЎЪЎМИЕНЫФЪЎМИЪБЃЌЮЂЖЏТіжБОЖНЋНЕЕЭ30%ЁЋ50%ЁЃгЩгкЮЂЖЏТіжБОЖВЛЛсЫцЯйпАХЈЖШЕФдіМѕЖјЮоЯодіМгЛђЧїгкСуЃЌЙЪЮвУЧЩшЖўепЕФЙиЯЕЮЊЃК (7)
(7)
, АйФДвНвЉ
ЦфжаЃКramaxКЭraminЮЊЮЂЖЏТіДњаЛЕїНкЕФзюДѓгызюаЁАыОЖЃЌcadoЮЊЯйпАЕФФГвЛВЮПМХЈЖШЁЃЬѕМўn>1ЪЧЮЊСЫЪЙЧњЯпГЪЁАSЁБаЮЃЌШчЭМ1ЫљЪОЁЃ
ЭМ1 ЮЂЖЏТіАыОЖгыЯйпАХЈЖШЕФЙиЯЕ
Fig 1 Relationship between arteriole radius and adenosine concentration
гЩгкМфжЪЯйпАХЈЖШИФБфЪБЃЌЪзЯШНЋв§Ц№УЋЯИбЊЙмУмЖШИФБфЃЌЖјЮЂЖЏТіЙмОЖЕФЯджјИФБфдђЗЂЩњдкУЋЯИбЊЙмУмЖШДяЕНзюДѓжЕжЎКѓЃЌЙЪЕквЛЕїНкСйНчХЈЖШCmlгІДѓгкЮЂЖЏТіЁАе§ГЃЁБХЈЖШCadnЧвЮЛгкЮЂЖЏТіАыОЖБфЛЏТЪНЯаЁЕФЮЛжУЁЃ
, http://www.100md.com
ЩЯЪНжаЕФСНИіГЃЪ§nКЭCadoПЩгЩвЛЖЈЕФвбжЊЬѕМўШЗЖЈЁЃИљОнKrollКЭStepp[16]ЕФЙРМЦЃЌЙЗаФМЁМфжЪЯйпАХЈЖШдМЮЊ100ЁЋ200 nMЁЃвдДЫзїЮЊЯйпАХЈЖШЕФЁАе§ГЃЁБжЕCadnЃЌдђПЩЕУЃК
Cad=CadnЪБЃК ra=ran
СэОнЧАЪіFrameКЭSarelius[13]ЕФбаОПНсЙћЃЌПЩЩшЯйпАХЈЖШЮЊCadM=10-4MЪБЃЌЮЂЖЏТіАыОЖвбЗЧГЃНгНќДњаЛЕїНкЕФМЋДѓжЕЃЌР§ШчЕШгк0.99ramaxЁЃетбљЃЌЮвУЧЕУЕНЃК
Cad=CadMЪБЃК ra=0.99ramax
, АйФДвНвЉ
НЋДЫЖўЪНДњШыЪН(7)ЃЌОЭПЩШЗЖЈГЃЪ§nКЭCadoЁЃгкЪЧЃЌЯйпАХЈЖШБфЛЏв§Ц№ЮЂЖЏТіШыПкАыОЖЕФИФБфПЩБэЮЊЃК

НЋМЁдадЕїПигыДњаЛадЕїПиНсКЯЦ№РДЃЌОЭЕУЕНЮЂЖЏТіАыОЖЕФОжВПЕїПиФЃаЭЃК (8)
(8)
ЦфжаЃК ЃЛPacЮЊЮЂЖЏТіЫњЯнЕФСйНчбЙСІЁЃ
ЃЛPacЮЊЮЂЖЏТіЫњЯнЕФСйНчбЙСІЁЃ
5 ЮЂЖЏТіздТЩдЫЖЏеёЗљЕФОжВППижЦФЃаЭ
баОПБэУїЃЌЮЂЖЏТібЊбЙдквЛЖЈЗЖЮЇФкБфЛЏЪБЃЌЖдздТЩдЫЖЏеёЗљУЛгагАЯьЃЛЖјЕБбЊбЙЕЭгкФГвЛСйНчжЕЪБЃЌздТЩдЫЖЏНЋЭЛШЛЭЃжЙЁЃгЩДЫЃЌЮвУЧПЩНЋздТЩдЫЖЏеёЗљЕФМЁдадПижЦБэЮЊЃК
, АйФДвНвЉ
b=bof(CNO).J(Pao-Pcrit) (9)
ЦфжаЃКboЮЊеёЗљГЃСПЃЛPcritЮЊздТЩдЫЖЏЭЛШЛЭЃжЙЕФСйНчбЙСІЃЛf(CNO)ЮЊвЛбѕЛЏЕЊЖдеёЗљЕФгАЯьКЏЪ§ЁЃгаЪЕбщБэУї[17]ЃЌИпЪГбЮдьГЩЕФИпбЊбЙ(166 mmHg)ФмЪЙЪѓЮЂЖЏТіздТЩдЫЖЏЕФеёЗљдіМгДя93%ЃЛзїепШЯЮЊЃЌЮЌГжздТЩдЫЖЏеёЗљВЛБфЕФЪЧЛљДЁвЛбѕЛЏЕЊ(Basal nitric oxide)ЃЌИпЪГбЮадИпбЊбЙаЮГЩЙ§ГЬжаЃЌвЛбѕЛЏЕЊЕФзїгУБЛвжжЦСЫЃЌЕМжТздТЩдЫЖЏеёЗљдіМгЁЃЮФЯз[18]вВБэУїЃЌвжжЦNOНЋдіМгЪѓФдЮЂЖЏТібЊСїЕФеёЗљЁЃвђДЫЃЌздТЩдЫЖЏеёЗљгІИУЪЧвЛбѕЛЏЕЊХЈЖШCNOЕФКЏЪ§ЁЃЕЋЪЧЃЌгЩгкДцдкЯрЗДЕФЪЕбщНсЙћЃЌМДдіМгNOФмдіЧПздТЩдЫЖЏЃЌМгжЎЖдЦфЛњжЦЛЙКмВЛСЫНтЃЌЙЪФПЧАЩаВЛФмИјГіКЏЪ§f(CNO)ЕФОпЬхаЮЪНЁЃ
, АйФДвНвЉ
6 Ьж Тл
ЮЂбЛЗОжВППижЦжївЊжИМЁдадПижЦгыДњаЛадПижЦЃЌгаЕФзїеп[19]дђШЯЮЊЛЙгІАќРЈФкЦЄЯИАћПижЦЁЃЮЂбЊЙмФкЦЄЯИАћБОЩэОпгаЕїНкЮЂбЛЗЕФЙІФмЃЌвбДгЕчОЕЕУЕНжЄЪЕЁЃФкЦЄЯИАћАћЬхЭЛЯђЧЛЯђ(гаЕФПЩДяЧЛФк1/2ЁЋ2/3)ЃЌбЊЙмБкОжВПАМЯнЕШЃЌЖМФмВЛЭЌГЬЖШЕигАЯьбЊСїЁЃвђДЫПЩвдШЯЮЊЃЌУЋЯИбЊЙмФкЦЄЯИАћЪЧЛ§МЋВЮгыЮЂбЛЗОжВПЕїНкЕФЁЃ
ДгзщжЏШБбѕЕНЮЂбЊЙмзїГіЗДгІгаМИУыЕНМИЪЎУыЕФжЭКѓЪБМф[14]ЁЃПЩвдШЯЮЊетОЭЪЧЯйпАДгЯИАћЕНУЋЯИбЊЙмЧАРЈдММЁЛђЮЂЖЏТіЕФдЫЖЏЪБМфЁЃгЩгкУЋЯИбЊЙмЧАРЈдММЁКЭЮЂЖЏТіДІЕФЯйпАХЈЖШВЛЖЯБфЛЏЃЌЪЙУЋЯИбЊЙмЗжВМгыЮЂЖЏТіЙмОЖВЛЖЯБфЛЏЃЌвђДЫЃЌзїЮЊМђЛЏЃЌЮвУЧШЁздТЩдЫЖЏЕФвЛИіжмЦкзїЮЊжЭКѓЪБМфЃЌвдвЛжмЦкФкФГЕуЯйпАХЈЖШЕФЦНОљжЕЪЧЗёГЌЙ§СйНчжЕзїЮЊв§Ц№ЕїНкЕФЬѕМўЃЌЖјЕїНкЕФНсЙћдђдкЯТвЛжмЦкФкБэЯжГіРДЁЃетвЛМђЛЏЕФИљОнЪЧЃЌздТЩдЫЖЏжмЦкдМ10 sзѓгвЃЌУЋЯИбЊЙмМфаЊадПЊБезДЬЌУПвЛЯрЕФбгајЪБМфдМ2ЁЋ8 sЃЌгыжЭКѓЪБМфНгНќЁЃ
, АйФДвНвЉ
гЩгкУЋЯИбЊЙмЧАРЈдММЁОрзщжЏШБбѕЧјНЯНќЃЌуажЕНЯЕЭЃЌЙЪЮЂбЛЗЯЕЭГЕквЛЕїНкзмБШЕкЖўЕїНкЯШЗЂЩњЁЃЪ§жЕМЦЫуНсЙћБэУїЃЌгЩгкзщжЏШБбѕЧјЗжВМдкОрУЋЯИбЊЙм(гШЦфЪЧЦфЮЂЖЏТіЖЫ)НЯдЖЕФЕиЗНЃЌЫљвдетаЉЕиЧјФкУЋЯИбЊЙмЕФПЊЗХЛђПЊЗХзХЕФУЋЯИбЊЙмЯђетаЉЕиЧјбгЩьЃЌЖдгкИФБфзщжЏШБбѕзДПіЃЌОпгазюживЊЕФвтвхЃЌФмдкВЛдіМгбЊСїСПЕФЧщПіЯТЃЌЯджјМѕаЁжБжСЭъШЋЯћУ№ШБбѕЧјЁЃШєУЋЯИбЊЙмЗжВМЗЖЮЇВЛБфЃЌНіУмЖШдіМгЃЌдђМфжЪбѕХЈЖШГ§ЗжВМгаЫљИФБфЭтЃЌЦНОљХЈЖШВЂЮоУїЯдБфЛЏЁЃетБэУїУЋЯИбЊЙмЁАЦНОљУмЖШЁБИХФюЕФгІгУЗЖЮЇЪЧгаЯоЕФЃЌКмЖрЪБКђЦфЗжВМзДПіЦ№зХживЊЕФзїгУЁЃМЦЫуНсЙћЛЙБэУїЃЌОрУЋЯИбЊЙмДѓгк100 ІЬmЕФЕиЗНЃЌдкздТЩдЫЖЏЕФећИіжмЦкФкЃЌМИКѕЖМДІгкШБбѕзДЬЌЁЃЮФЯз[20]жИГіЃЌе§ГЃШЫЬхЯИАћгыУЋЯИбЊЙмЕФОрРыаЁгк50 ІЬmЁЃ
ШБбѕЧјЯћУ№КѓЃЌМфжЪЯйпАХЈЖШНЕЕЭЃЌВПЗжУЋЯИбЊЙмЧАРЈдММЁжиаТЙиБеЁЃВЛЩйЪЕбщЙлВтЖМЗЂЯждке§ГЃЩњРэЬѕМўЯТЃЌУЋЯИбЊЙмФкЕФСїЖЏЪБЖЯЪБајЃЌДѓаЁКЭЗНЯђЖМОГЃИФБфЁЃгаЕФбаОПеп[21]ИќЗЂЯжЫљЙлВтЕФШЋВПУЋЯИбЊЙмОљгаГЄЖЬВЛЕШЕФСуСїСПЪБМфЃЌВЂШЯЮЊетЪЧгЩWBCЖТШћдьГЩЕФЁЃЮвУЧЕФбаОПдђБэУїЃЌУЋЯИбЊЙмЕквЛЕїНквВПЩвддьГЩетвЛЯжЯѓЁЃ
, АйФДвНвЉ
ЙњМвздШЛПЦбЇЛљН№зЪжњЯюФП(19472067)
ВЮПМЮФЯз
1ЃЌЮтЦфЯФжїБр.ЬхвКвђЫиКЭбЊвКбЛЗВЁРэЩњРэбЇ.ББОЉЃКББОЉвНПЦДѓбЇЁЂжаЙњаКЭвНПЦДѓбЇСЊКЯГіАцЩчЃЌ1991
2ЃЌGranger HJ,Borders JLЃЌMeininger GA et al.Microcirculatory control systems. in: The physiology and pharmacology of the microcirculation, vol.1, Mortillaro, NA(ed), Academic Press,1983ЁУ209
3ЃЌGranger HJЃЌShepherd AP. Dynamics and control of the microcirculation. Adv.in Biomed. Eng.1979;7ЁУ1
, АйФДвНвЉ
4ЃЌTakahashi EЃЌDoi K. Regulation of oxygen diffusion in hypoxic isolated cardiac myocytes. Am. J. Physiol,1996ЃЛ271ЁУH1734
5ЃЌHoshi Y, Hazeki OЃЛTamura M. Oxygen dependence of redox state of copper in cytochrome oxydase in vitro. J.Appl.Physiol.1993;74ЁУ1622
6ЃЌHester RL. Uptake of metabolites by postcapillary venules: Mechanism for the control of arteriolar diameter. Microvasc. Res,1993;46(2)ЁУ254
7ЃЌDietrick HHЃЌTyml K. Capillary as a communicating medium in the microvasculature. Microvasc. Res.1992ЃЛ43(1)ЁУ87
, АйФДвНвЉ
8ЃЌMitchell DЃЌYu KЃЌTyml K. Comparable effects of arteriolar and capillary stimuli on blood flow in rat skeletal muscle. Microvasc. Res.1997;53(1)ЁУ22
9ЃЌBertuglia S,Colantuoni AЃЌArnold M et al.Dynamic coherence analysis of vasomotion and flow motion in skeletal muscle microcirculation. Microvasc. Res,1996;52(3)ЁУ235
10ЃЌDuran WN,Mortimer PNЃЌLemich JM et al.Capillary reserve in isometrically contracting dog hearts. Am J. Physiol.1977ЃЛ233ЁУH276
, АйФДвНвЉ
11ЃЌAschheim E.Passage of substances across the walls of blood vessels: kinetics and mechanism. in: Microcirculation. vol.1,Kaley, G et al.(eds).Univ.Park Press. Baltimore, London,Tokyo,1977ЁУ213
12ЃЌLee SЃЌSchmid-Schonbein GW. Biomechanical model for the myogenic response in the microcirculation:Part I-Formulation and initial testing. J.Biomech.Eng.1996;118ЁУ145
13ЃЌFrame MDSЃЌSarelius IH. Arteriolar bifurcation angles vary with position and when flow is changed. Microvasc.Res,1993;46(2)ЁУ190
, http://www.100md.com
14ЃЌKontos HA,Fingar MHЃЌWieman JY et al.Role of tissue hypoxia in local regulation of cerebrol microcirculation. Am. J. Physiol,1978;234ЁУH582
15ЃЌKunert MP, Liard JFЃЌAbraham DJ et al.Low-affinity hemogl-
obin increases tissue PO2 and decreases arteriolar diameter and flow in the rat cremaster muscle. Microvasc. Res.1996;52(1)ЁУ58
16ЃЌKroll KЃЌStepp DW. Adenosine kinetics in canine coronary circulation. Am J. Physiol.1996;270(4)ЁУH1469
, http://www.100md.com
17ЃЌBoegehold MA. Enhanced arteriolar vasomotion in rats with chronic salt-induced hypertension. Microvasc.Res.1993;45(1)ЁУ83
18ЃЌDirnagl U, Lindauer UЃЌVillringer A. Nitric oxide synthase blockade enhances vasomotion in the cerebral microcirculation of anesthetized rats. Microvasc.Res.1993;45(3)ЁУ318
19ЃЌЬя ХЃ.ЮЂбЛЗ.ББОЉЃКПЦбЇГіАцЩчЃЌ1986ЁУ1
20ЃЌЮфаЧЛЇБр.ШЫЬхе§ГЃЩњРэЪ§Он.ББОЉЃКПЦбЇЦеМАГіАцЩчЃЌ1987ЁУ55
21ЃЌMayrovitz MN. Skin capillary matrics and hemodynamics in the hairless mouse. Microvasc.Res,1992;43(1)ЁУ46
(ЪеИхЃК1998-12-17), АйФДвНвЉ
ЕЅЮЛЃКЙљжйШ§(жаЙњПЦбЇдК ГЩЖММЦЫуЛњгІгУбаОПЫљздЖЏЭЦРэЪЕбщЪвЃЌГЩЖМ 610041)
ЙиМќДЪЃКЮЂбЛЗЃЛОжВППижЦЃЛЩњЮяСІбЇФЃаЭ
ЩњЮявНбЇЙЄГЬбЇдгжО000204 еЊвЊ ЗжЮіСЫМфжЪФкбЊЙмДњаЛЮяжЪЕФВњЩњКЭдЫЖЏЁЂУЋЯИбЊЙмЧАРЈдММЁЕФПЊБеЙ§ГЬЁЂЮЂЖЏТіЦНОљАыОЖЕФгАЯьвђЫиМАЮЂЖЏТіздТЩдЫЖЏеёЗљЕФБфЛЏЧщПіЃЌНЈСЂСЫЮЂбЛЗЯЕЭГОжВППижЦЙ§ГЬЕФЩњЮяСІбЇФЃаЭЃЌгажњгкЖдИУЙ§ГЬНјааЖЈСПЗжЮіЁЃ
A Biomechanical Model of Local Control of Microcirculation
Guo Zhongsan
(Chengdu Institute of Computer Application, Academia Sinica, Chengdu 610041)
, http://www.100md.com
Abstract In this paper we analyze the generation and motion of vasoactive metabolites in interstitium, the open and closure of precapillary sphincters, the factors affecting mean arteriolar diameter, and the change of amplitude of arteriolar vasomotion. A biomechanical model of local control of microcirculation has been developed. It is useful in quantitative analysis of microcirculatory local control.
Key words Microcirculation Local control Biomechanical model
, АйФДвНвЉ
1 в§ бд
ЮЂбЊЙмДВЪЧзщжЏЙрСїЁЂбЊвК-зщжЏСїЬхНЛЛЛКЭзщжЏбЊШнСПЕФПижЦГЁЫљЁЃжївЊЕФПижЦепЪЧЮЂЖЏТіКЭУЋЯИбЊЙмЧАРЈдММЁЃЌжївЊПижЦЛњжЦгаОжВППижЦ(Local control,ЛђIntrinsic control)КЭШЋЩэПижЦ(Remote control,ЛђExtrinsic control)ЃЌжївЊПижЦФПБъЪЧдкИїжжЛЗОГЬѕМўИФБф(зюГЃМћЕФЪЧбЊбЙКЭбѕХЈЖШИФЖШ)ЪББЃГжФкЛЗОГЕФЮШЖЈадЁЃ
ШЋЩэПижЦжївЊжИЩёОПижЦгыЬхвКПижЦЁЃЖўепЙиЯЕЪЎЗжУмЧаЃЌвђЮЊЩёОПижЦвВЪЧЭЈЙ§ЬхвКвђзгЪЕЯжЕФ[1]ЃЌЬхвКвђзгЕФДѓВПЗжЪЧЩёОЕнжЪЃЌШчШЅМзЩіЩЯЯйЫиЁЂввѕЃЕЈМюЕШЃЌгаЕФдђЪЧЯИАћЕФЪЭЗХВњЮяЃЌШчЯйпА(adenosine)ЁЃ
ОжВППижЦАќРЈМЁдадПижЦгыДњаЛадПижЦЁЃМЁдадПижЦЪЧжИЮЂЖЏТідкФкЭтбЙВюдіДѓЪБжїЖЏЪеЫѕвдПижЦбЊСїСПЕФЯжЯѓЁЃДњаЛадПижЦЪЧЭЈЙ§ЯИАћДњаЛВњЮяНјааЕФПижЦЁЃгабаОПШЯЮЊ[1]ЃЌЮЂбЛЗЕФОжВПЕїНкжаЃЌ75%ЪЧДњаЛадЕїНкЁЃ
, http://www.100md.com
ЮЂбЛЗЯЕЭГПижЦЙ§ГЬЕФЖЈСПбаОПЙЄзїЗЧГЃЩйЁЃзїепЗжЮібаОПСЫгаЙиЕФЩњРэЙ§ГЬЃЌНЈСЂСЫЮЂбЛЗЯЕЭГОжВППижЦЙ§ГЬЕФЩњЮяСІбЇФЃаЭЃЌгаРћгкЖдетвЛЙ§ГЬНјааЖЈСПЗжЮіЁЃ
2 МфжЪЯйпАдДЧПЖШМАЯйпАХЈЖШЗжВМЙЋЪН
ЯИАћ(МйЩшОљдШЗжВМдкМфжЪПеМфФк)ЪЭЗХЛюбЊЙмДњаЛЮяжЪ(дкаФМЁжаЃЌзюживЊЕФЪЧЯйпАЃЛЮЊа№ЪіЗНБуЃЌБОЮФжаЮвУЧгУЯйпАДњБэЫљгаПЩФмЕФЛюбЊЙмДњаЛЮяжЪ)ШЁОігкЯпСЃЬхЕФбѕРћгУТЪЁЃЯпСЃЬхФкЕФЯИАћЩЋЫи(Cytochrom)aa3вдЛЙдгыбѕЛЏСНжжЗНЪНДцдкВЂПЩЛЅЯрзЊЛЏЁЃбѕНјШыЯИАћКѓЃЌМДзїЮЊЕчзгЕФЪмЬхгызїЮЊЕчзгЙЉЬхЕФЛЙдaa3ЯрЛЅзїгУЃЌЖјЯпСЃЬхЕФбѕРћгУТЪUO2MMОЭгЩЯИАћФкЕчзгЪмЬхЕФХЈЖШCO2cellcellгыЕчзгЙЉЬхЕФХЈЖШ[aa3]RЙВЭЌОіЖЈ[2]ЃК
(1), АйФДвНвЉ
дкОВЯЂзДЬЌЯТЃЌ[aa3]RжЛеМЯИАћЩЋЫизмХЈЖШ[aa3]TЕФ1/10ЛђИќЩйЁЃЕБЯИАћбѕХЈЖШНЕЕЭЪБЃЌбѕЛЏзїгУМѕШѕЃЌaa3ЕФбѕЛЏ-ЛЙдЖЏЬЌЦНКтБЛЦЦЛЕЃЌВПЗжбѕЛЏЕФaa3БЛЛЙдЃЌ[aa3]RдіМгЃЌДгЖјВЙГЅСЫCO2cellcellНЕЕЭЕФгАЯьЃЌЪЙUO2MMЮЌГжВЛБфЛђЛљБОВЛБфЁЃетжжВЙГЅзїгУНјааЕНaa3ШЋВПЛЙдЪБЃЌЯИАћбѕХЈЖШОЭЕНСЫвЛИіСйНчжЕCO2ccccЃЌНјвЛВННЕЕЭОЭЛсЪЙUO2MMЯджјМѕаЁЃЌЯИАћПЊЪМДѓСПЪЭЗХЯйпАЁЃОнДЫПЩЕУ[2]ЃК
(2), АйФДвНвЉ
ЦфжаЃКPO2ЊЊcellЮЊЯИАћбѕЗжбЙЃЛUO2ЊЊmaxЮЊЯпСЃЬхзюДѓбѕРћгУТЪЃЌГЃСПЃЛKMЮЊЯИАћбѕЗжбЙВЮПМжЕЃЌБэЪОЯпСЃЬхбѕРћгУТЪЮЊзюДѓбѕРћгУТЪЕФ50%ЪБЕФЯИАћбѕЗжбЙЁЃдкДѓЖрЪ§зщжЏжаЃЌKM<0.1 mmHgЁЃЖјЭЈГЃЯИАћбѕЗжбЙБШKMДѓ1ЁЋ2ИіЪ§СПМЖ[37ЁцЪБдМЮЊ2ЁЋ5 mmHg(1mmHg=0.133kPa)]ЁЃвђДЫЃЌЯИАћбѕХЈЖШдкЯрЕБДѓЕФЗЖЮЇФкБфЛЏЪБЃЌUO2ЊЊMЕФБфЛЏЖМЗЧГЃаЁЁЃвЛАуЕиЫЕЃЌЯИАћбѕЗжбЙДѓгк1 mmHgЪБЃЌОљПЩНќЫЦШЯЮЊUO2ЊЊMЪЧвЛИіГЃСПЁЃдкБОЮФжаЃЌЮвУЧгУЯпСЃЬхЯрЖдбѕРћгУТЪ
, http://www.100md.com
БэЪОЯИАћЪЧЗёШБбѕМАШБбѕГЬЖШЃЛР§ШчЃЌUO2rr<0.95ЮЊЧсЖШШБбѕЃЌUO2rr<0.90ЮЊжиЖШШБбѕЃЌЕШЕШЁЃ
ЯИАћбѕХЈЖШгЩЯТЪНИјГі[3]ЃК
ЦфжаЃКVcellЮЊЯИАћЬхЛ§ЃЌJO2cellcellЮЊНјШыЯИАћЕФбѕЭЈСПЃК
ЦфжаЃКCO2IIЮЊМфжЪбѕХЈЖШЁЃгЩгкЖдбѕЗжзгРЉЩЂЕФзшСІЃЌЯИАћФкЭтДцдкбѕХЈЖШЬнЖШЁЃЪЕбщБэУїЃЌЯИАћОпгаЕїНкетвЛбѕХЈЖШЕФФмСІЃЌдкМфжЪШБбѕЪБМѕаЁетвЛЬнЖШЃЌДгЖјВПЗжЕиЮЌГжЯИАће§ГЃЕФбѕКЯзїгУ[4]ЁЃетжжЙІФмадБфЛЏПЩФмЪЧвђЮЊaa3гыбѕЕФЧзКЯСІЫцЯИАћЕФФмСПзДПіЗЂЩњИФБфЫљв§Ц№ЕФ[5]ЁЃвђДЫЃЌЩЯЪНжаЕФЯЕЪ§Kc2гІИУЪЧ[aa3]RЕФКЏЪ§ЁЃЕЋдкБОЮФжаЃЌЮвУЧднШЁKc2ЮЊГЃСПЁЃ
, http://www.100md.com
ПЩвдПДГіЃЌЯпСЃЬхбѕРћгУТЪЁЂЯИАћбѕХЈЖШКЭМфжЪбѕХЈЖШЕФЙиЯЕЪЧКмИДдгЕФЁЃзїЮЊМђЛЏЃЌЮвУЧМйЩшЯИАћбѕХЈЖШЫцЪБМфБфЛЏКмТ§ЃЌНјШыЯИАћЕФбѕгыЯпСЃЬхЯћКФЕФбѕЫцЪББЃГжЦНКтЃЌДгЖјЕУЕНЃК
ЦфжаЃК
ЮЊЖдгІгкKMЕФбѕХЈЖШЁЃИљОнЩЯЪіЗжЮіЃЌВЂМйЩшЯИАћЪЭЗХЯйпАЕФЧПЖШгыЯпСЃЬхбѕРћгУТЪГЩЯпадЙиЯЕЃЌЮвУЧЕУЕНШчЯТЯйпАдДЧПЖШЙЋЪНЃК
(3)ЦфжаЃКSadЮЊМфжЪЯйпАдДЧПЖШЃЛKadЮЊЯйпАдДЧПЖШЯЕЪ§ЁЃ
ДгЯйпАдДВњЩњЕФЯйпАдкМфжЪФкЕФЪфдЫЙ§ГЬгыбѕЯрЫЦЃЌвВАќРЈРЉЩЂЁЂЖдСїКЭЛњаЕУжЩЂШ§ЗНУцЃЌЧвврПЩМйЩшМфжЪЖдЯйпАЮоХХГтзїгУЁЃДЫЭтЃЌбЊвКбЛЗКЭУЋЯИбЊЙмЭјЕФДцдкЖдЯйпАЕФЗжВМКЭдЫЖЏФмВњЩњКмДѓгАЯьЁЃЪЕбщБэУїЃЌдкЮЂОВТізЂШы0.1 ІЬMХЈЖШЕФЯйпАЃЌФмЯджјРЉеХЯрСкЮЂЖЏТіЕФЙмОЖ[6]ЃЌЫЕУїдкЮЂЖЏТі-ЮЂОВТіжЎМфДцдкЁАРЉЩЂНЛСЌЁБЃЌЪЙвбОЁАРыПЊЁБСЫЮЂбЛЗЕЅдЊЕФЯйпАШдФмВњЩњгАЯьЁЃдкУЋЯИбЊЙмФкзЂШыШЅМзЩіЩЯЯйЫиЃЌФмЪЙЙЉбЊЮЂЖЏТіЙмОЖМѕаЁЃЌУЋЯИбЊЙмСїЫйНЕЕЭ(жБжСНЕЮЊСу)ЃЛзЂШыввѕЃЕЈМяЛђЛКМЄыФЃЌПЩЪЙЮЂЖЏТіРЉеХВЂдіМгбЊСїЫйЖШ[7ЃЌ8]ЁЃзїепШЯЮЊЃЌетжжДгУЋЯИбЊЙмЕНЮЂЖЏТіЕФЭЫааадЕїНкЃЌМШВЛЪЧДгУЋЯИбЊЙмжБНгЯђЮЂЖЏТіРЉЩЂЕФНсЙћЃЌвВВЛЪЧЭЈЙ§ЮЂЖЏТі-ЮЂОВТіРЉЩЂНЛСЌЪЕЯжЕФЃЌЖјЪЧвђЮЊУЋЯИбЊЙмБОЩэОЭОпгаДЋЪфЛюбЊЙмЮяжЪЕФЙІФмЁЃДЫЭтЃЌУЋЯИбЊЙмФкЦЄЯИАћЛЙФмЮќЪеДѓСПЯйпА[9]ЁЃвђДЫЃЌЛюбЊЙмЮяжЪдкЮЂбЛЗЕЅдЊжаЕФдЫЖЏЗНЪНПЩФмЪЎЗжИДдгЃЌгаД§НјвЛВНбаОПЁЃзїЮЊМђЛЏЃЌзїепгУдДЕуЕќМгЗЈМЦЫуМфжЪИїДІЕФЯйпАХЈЖШЃЌiЕуЕФЯйпАдДSadiдкjЕуВњЩњЕФЯйпАХЈЖШCadI(ij)гЩЦНЗНЗДБШТЩИјГіЃК
, http://www.100md.com
ЦфжаЃКXijЮЊiЁЂjжЎМфЕФОрРыЁЃетбљЃЌФГвЛЪБПЬjЕуЕФЯйпАХЈЖШОЭгЩЯТЪНИјГіЃК
(4)ЦфжаЃКCadI(j)ЮЊМфжЪФкФГЕуjЕФЯйпАХЈЖШЃЛSadiЮЊФГЕуiЕФЯйпАдДЧПЖШЁЃЦфЛ§ЗжаЮЪНЮЊЃК
ЦфжаЃКVЮЊМфжЪЬхЛ§ЁЃ
3 УЋЯИбЊЙмЧАРЉдММЁЕФОжВППижЦФЃаЭ
НќДњЕФбаОПБэУї[10ЃЌ11]ЃЌУЋЯИбЊЙмЕФДЂБИ(Reserve)гыФММЏ(Recruitment)ЬсЙЉСЫвЛИіОЋЧЩЕФЕїНкЛњжЦЃЛзщжЏФкЙиБезХЕФУЋЯИбЊЙмашвЊЪБПЩвдПЊЗХЃЌПЊЗХзХЕФУЋЯИбЊЙмврПЩФмЙиБеЃЌЪЙбЊвКдкЮЂбЊЙмДВФкИќгааЇЕижиаТЗжВМЃЌДгЖјдкбЊСїСПВЛБфЕФЬѕМўЯТБуПЩТњзудіМгСЫЕФашвЊЁЃетжжЕїНкЪЧЮЂбЛЗЕЅдЊФкВПЕФЕїНкЃЌВЛгАЯьЦфгрЕЅдЊЕФЙрСїЃЌВЛдіМгаФдрБУбЊЙІФмЃЌдкБОЮФжаЮвУЧГЦжЎЮЊФкВПЕїНкЛђЕквЛЕїНкЁЃ
, http://www.100md.com
ЕБбЊвКЗжВМдкЫљгаПЩНЛЛЛУцЩЯШдВЛФмТњзузщжЏЕФашвЊЪБЃЌЮЂЖЏТіНЋЛсЯджјЕиРЉеХЃЌЪЙНјШыИУЮЂбЛЗЕЅдЊЕФбЊСїСПдіМгЃЌДгЖјТњзузщжЏЕФашвЊЁЃетжжЕїНкЗНЪНЛсИФБфЦфгрЮЂбЛЗЕЅдЊЕФбЊСїзДПіЃЌЛђдіМгаФдрИККЩЃЌвђДЫЮвУЧГЦжЎЮЊЭтВПЕїНкЛђЕкЖўЕїНкЁЃ
ЩшУЋЯИбЊЙмЧАРЈдММЁДІЕФЯйпАХЈЖШДѓгкЕквЛЕїНкСйНчжЕCmlЪБЃЌИУУЋЯИбЊЙмМДЭъШЋПЊЗХЃЌЗДжЎдђЭъШЋЙиБеЃЌБуЕУЕНШчЯТЕФУЋЯИбЊЙмЧАРЈдММЁДњаЛадЕїПиФЃаЭЃК
(5)ЦфжаЃКrЊcЮЊУЋЯИбЊЙмАыОЖЃЛrЊcoЮЊАыОЖГЃСПЃЛJ(x)ЮЊНздОКЏЪ§ЁЃ
4 ЮЂЖЏТіЦНОљАыОЖЕФОжВППижЦФЃаЭ
ЮЂЖЏТіМЁдадПижЦЕФЛњРэЛЙВЛКмЧхГўЃЌФПЧАЩаЮоЦеБщНгЪмЕФЩњЮяСІбЇФЃаЭЁЃЪЕМЪгІгУжаГЃВЩгУLaplaceЙЋЪНЃК
, http://www.100md.com
rp=ГЃЪ§
ЦфжаЃКrЮЊЮЂЖЏТіАыОЖЃЛpЮЊбЙСІЁЃетЪЧИљОнЮЂЖЏТіБкФкКЯСІВЛБфЕФМйЩшЖјЕУЕНЕФЁЃгЩгкЮЂЖЏТіЪєКёБкЙмЃЌЙЪЮвУЧМйЩшМЁдадЕїНкЕФФПЕФЪЧБЃГжЮЂЖЏТіБкФкЦНОљгІСІІвaВЛБфЃЌМДЃК
rp=Івah hЁЊБкКё
ЭЌЪБМйЩшЙмБкВЛПЩбЙЫѕЃЌгЩДЫЕУЕНЮЂЖЏТіЕФМЁдадЕїПиФЃаЭЃК
(6)ЦфжаЃКa0ЮЊЮЂЖЏТіШыПкАыОЖЦНОљжЕЃЛa00ЮЊе§ГЃЧщПіЯТЮЂЖЏТіШыПкАыОЖЦНОљжЕЃЛPa0ЮЊЮЂЖЏТіШыПкДІбЊбЙЃЛPЁфaoЮЊе§ГЃЧщПіЯТЮЂЖЏТіШыПкДІбЊбЙЃЛ
ЃЛh0ЮЊЮЂЖЏТіШыПкДІЙмБкКёЖШЁЃгЩгкЪНжаАќКЌСЫАыОЖгыБкКёжЎБШЃЌЙЪБШLaplaceЙЋЪНИќФмЗДгГВЛЭЌЮЂЖЏТіЕФВюБ№ЁЃПЩвдПДГіЃЌІвaдНаЁЃЌЩЯЪНдННгНќгкLaplaceЙЋЪНЁЃ, АйФДвНвЉ
ДгВЛЭЌЕФНЧЖШПЩвдИјГіВЛЭЌЕФФЃаЭЁЃР§ШчЃЌгаЕФзїеп[12]ЭЈЙ§в§ШыЁАВЮПМжБОЖЁБЕФИХФюРДНЈСЂЮЂЖЏТіМЁдадЕїПиФЃаЭЁЃжЕЕУжИГіЕФЪЧЃЌИУЮФзїепЕУЕНЕФrЫцPЕФБфЛЏЫйЖШКЭДЫДІЕФ(6)ЪНЖМБШLaplaceЙЋЪНЫљИјГіЕФЫйЖШТ§ЁЃ
ЮЂЖЏТіАыОЖЫцЮЂЖЏТіШыПкДІЕФЯйпАХЈЖШдіМѕЖјдіМѕЁЃИљОнЖдН№ЛЦВжЪѓЕФЪЕбщ[13]ЃЌЕБЯйпАХЈЖШЮЊ10-4MзѓгвЪБЃЌЮЂЖЏТіРЉеХДяЕНзюДѓжЕЁЃИїМЖЮЂЖЏТіЕФОВЯЂжБОЖдМЮЊзюДѓжБОЖЕФ70%ЁЃЕБзщжЏбѕХЈЖШГЌЙ§ашвЊЪБЃЌЮЂЖЏТіЙмОЖНЋМѕаЁЃЌДгЖјЬсЙЉвЛИіИКЗДРЁЕїНкЃЌЪЙзщжЏбѕЙЉгІгыЯћКФЧїгкЦНКтЁЃЕЋдкЖЈСПбаОПЩЯФПЧАДцдкНЯДѓВюБ№ЃКгаЕФШЯЮЊЖЏТібѕЗжбЙИпгке§ГЃжЕЪБЃЌЮЂЖЏТіжБОЖНігаКмаЁЕФМѕЩй[14]ЃЌЖјKunertЕШ[15]гУНЕЕЭбЊЧђбѕЧзКЭСІЕФЗНЗЈЕУЕНЕФНсЙћдђЪЧЕБзщжЏбѕЗжбЙдіМгАйЗжжЎЪЎМИЕНЫФЪЎМИЪБЃЌЮЂЖЏТіжБОЖНЋНЕЕЭ30%ЁЋ50%ЁЃгЩгкЮЂЖЏТіжБОЖВЛЛсЫцЯйпАХЈЖШЕФдіМѕЖјЮоЯодіМгЛђЧїгкСуЃЌЙЪЮвУЧЩшЖўепЕФЙиЯЕЮЊЃК
(7), АйФДвНвЉ
ЦфжаЃКramaxКЭraminЮЊЮЂЖЏТіДњаЛЕїНкЕФзюДѓгызюаЁАыОЖЃЌcadoЮЊЯйпАЕФФГвЛВЮПМХЈЖШЁЃЬѕМўn>1ЪЧЮЊСЫЪЙЧњЯпГЪЁАSЁБаЮЃЌШчЭМ1ЫљЪОЁЃ
ЭМ1 ЮЂЖЏТіАыОЖгыЯйпАХЈЖШЕФЙиЯЕ
Fig 1 Relationship between arteriole radius and adenosine concentration
гЩгкМфжЪЯйпАХЈЖШИФБфЪБЃЌЪзЯШНЋв§Ц№УЋЯИбЊЙмУмЖШИФБфЃЌЖјЮЂЖЏТіЙмОЖЕФЯджјИФБфдђЗЂЩњдкУЋЯИбЊЙмУмЖШДяЕНзюДѓжЕжЎКѓЃЌЙЪЕквЛЕїНкСйНчХЈЖШCmlгІДѓгкЮЂЖЏТіЁАе§ГЃЁБХЈЖШCadnЧвЮЛгкЮЂЖЏТіАыОЖБфЛЏТЪНЯаЁЕФЮЛжУЁЃ
, http://www.100md.com
ЩЯЪНжаЕФСНИіГЃЪ§nКЭCadoПЩгЩвЛЖЈЕФвбжЊЬѕМўШЗЖЈЁЃИљОнKrollКЭStepp[16]ЕФЙРМЦЃЌЙЗаФМЁМфжЪЯйпАХЈЖШдМЮЊ100ЁЋ200 nMЁЃвдДЫзїЮЊЯйпАХЈЖШЕФЁАе§ГЃЁБжЕCadnЃЌдђПЩЕУЃК
Cad=CadnЪБЃК ra=ran
СэОнЧАЪіFrameКЭSarelius[13]ЕФбаОПНсЙћЃЌПЩЩшЯйпАХЈЖШЮЊCadM=10-4MЪБЃЌЮЂЖЏТіАыОЖвбЗЧГЃНгНќДњаЛЕїНкЕФМЋДѓжЕЃЌР§ШчЕШгк0.99ramaxЁЃетбљЃЌЮвУЧЕУЕНЃК
Cad=CadMЪБЃК ra=0.99ramax
, АйФДвНвЉ
НЋДЫЖўЪНДњШыЪН(7)ЃЌОЭПЩШЗЖЈГЃЪ§nКЭCadoЁЃгкЪЧЃЌЯйпАХЈЖШБфЛЏв§Ц№ЮЂЖЏТіШыПкАыОЖЕФИФБфПЩБэЮЊЃК
НЋМЁдадЕїПигыДњаЛадЕїПиНсКЯЦ№РДЃЌОЭЕУЕНЮЂЖЏТіАыОЖЕФОжВПЕїПиФЃаЭЃК
(8)ЦфжаЃК
ЃЛPacЮЊЮЂЖЏТіЫњЯнЕФСйНчбЙСІЁЃ5 ЮЂЖЏТіздТЩдЫЖЏеёЗљЕФОжВППижЦФЃаЭ
баОПБэУїЃЌЮЂЖЏТібЊбЙдквЛЖЈЗЖЮЇФкБфЛЏЪБЃЌЖдздТЩдЫЖЏеёЗљУЛгагАЯьЃЛЖјЕБбЊбЙЕЭгкФГвЛСйНчжЕЪБЃЌздТЩдЫЖЏНЋЭЛШЛЭЃжЙЁЃгЩДЫЃЌЮвУЧПЩНЋздТЩдЫЖЏеёЗљЕФМЁдадПижЦБэЮЊЃК
, АйФДвНвЉ
b=bof(CNO).J(Pao-Pcrit) (9)
ЦфжаЃКboЮЊеёЗљГЃСПЃЛPcritЮЊздТЩдЫЖЏЭЛШЛЭЃжЙЕФСйНчбЙСІЃЛf(CNO)ЮЊвЛбѕЛЏЕЊЖдеёЗљЕФгАЯьКЏЪ§ЁЃгаЪЕбщБэУї[17]ЃЌИпЪГбЮдьГЩЕФИпбЊбЙ(166 mmHg)ФмЪЙЪѓЮЂЖЏТіздТЩдЫЖЏЕФеёЗљдіМгДя93%ЃЛзїепШЯЮЊЃЌЮЌГжздТЩдЫЖЏеёЗљВЛБфЕФЪЧЛљДЁвЛбѕЛЏЕЊ(Basal nitric oxide)ЃЌИпЪГбЮадИпбЊбЙаЮГЩЙ§ГЬжаЃЌвЛбѕЛЏЕЊЕФзїгУБЛвжжЦСЫЃЌЕМжТздТЩдЫЖЏеёЗљдіМгЁЃЮФЯз[18]вВБэУїЃЌвжжЦNOНЋдіМгЪѓФдЮЂЖЏТібЊСїЕФеёЗљЁЃвђДЫЃЌздТЩдЫЖЏеёЗљгІИУЪЧвЛбѕЛЏЕЊХЈЖШCNOЕФКЏЪ§ЁЃЕЋЪЧЃЌгЩгкДцдкЯрЗДЕФЪЕбщНсЙћЃЌМДдіМгNOФмдіЧПздТЩдЫЖЏЃЌМгжЎЖдЦфЛњжЦЛЙКмВЛСЫНтЃЌЙЪФПЧАЩаВЛФмИјГіКЏЪ§f(CNO)ЕФОпЬхаЮЪНЁЃ
, АйФДвНвЉ
6 Ьж Тл
ЮЂбЛЗОжВППижЦжївЊжИМЁдадПижЦгыДњаЛадПижЦЃЌгаЕФзїеп[19]дђШЯЮЊЛЙгІАќРЈФкЦЄЯИАћПижЦЁЃЮЂбЊЙмФкЦЄЯИАћБОЩэОпгаЕїНкЮЂбЛЗЕФЙІФмЃЌвбДгЕчОЕЕУЕНжЄЪЕЁЃФкЦЄЯИАћАћЬхЭЛЯђЧЛЯђ(гаЕФПЩДяЧЛФк1/2ЁЋ2/3)ЃЌбЊЙмБкОжВПАМЯнЕШЃЌЖМФмВЛЭЌГЬЖШЕигАЯьбЊСїЁЃвђДЫПЩвдШЯЮЊЃЌУЋЯИбЊЙмФкЦЄЯИАћЪЧЛ§МЋВЮгыЮЂбЛЗОжВПЕїНкЕФЁЃ
ДгзщжЏШБбѕЕНЮЂбЊЙмзїГіЗДгІгаМИУыЕНМИЪЎУыЕФжЭКѓЪБМф[14]ЁЃПЩвдШЯЮЊетОЭЪЧЯйпАДгЯИАћЕНУЋЯИбЊЙмЧАРЈдММЁЛђЮЂЖЏТіЕФдЫЖЏЪБМфЁЃгЩгкУЋЯИбЊЙмЧАРЈдММЁКЭЮЂЖЏТіДІЕФЯйпАХЈЖШВЛЖЯБфЛЏЃЌЪЙУЋЯИбЊЙмЗжВМгыЮЂЖЏТіЙмОЖВЛЖЯБфЛЏЃЌвђДЫЃЌзїЮЊМђЛЏЃЌЮвУЧШЁздТЩдЫЖЏЕФвЛИіжмЦкзїЮЊжЭКѓЪБМфЃЌвдвЛжмЦкФкФГЕуЯйпАХЈЖШЕФЦНОљжЕЪЧЗёГЌЙ§СйНчжЕзїЮЊв§Ц№ЕїНкЕФЬѕМўЃЌЖјЕїНкЕФНсЙћдђдкЯТвЛжмЦкФкБэЯжГіРДЁЃетвЛМђЛЏЕФИљОнЪЧЃЌздТЩдЫЖЏжмЦкдМ10 sзѓгвЃЌУЋЯИбЊЙмМфаЊадПЊБезДЬЌУПвЛЯрЕФбгајЪБМфдМ2ЁЋ8 sЃЌгыжЭКѓЪБМфНгНќЁЃ
, АйФДвНвЉ
гЩгкУЋЯИбЊЙмЧАРЈдММЁОрзщжЏШБбѕЧјНЯНќЃЌуажЕНЯЕЭЃЌЙЪЮЂбЛЗЯЕЭГЕквЛЕїНкзмБШЕкЖўЕїНкЯШЗЂЩњЁЃЪ§жЕМЦЫуНсЙћБэУїЃЌгЩгкзщжЏШБбѕЧјЗжВМдкОрУЋЯИбЊЙм(гШЦфЪЧЦфЮЂЖЏТіЖЫ)НЯдЖЕФЕиЗНЃЌЫљвдетаЉЕиЧјФкУЋЯИбЊЙмЕФПЊЗХЛђПЊЗХзХЕФУЋЯИбЊЙмЯђетаЉЕиЧјбгЩьЃЌЖдгкИФБфзщжЏШБбѕзДПіЃЌОпгазюживЊЕФвтвхЃЌФмдкВЛдіМгбЊСїСПЕФЧщПіЯТЃЌЯджјМѕаЁжБжСЭъШЋЯћУ№ШБбѕЧјЁЃШєУЋЯИбЊЙмЗжВМЗЖЮЇВЛБфЃЌНіУмЖШдіМгЃЌдђМфжЪбѕХЈЖШГ§ЗжВМгаЫљИФБфЭтЃЌЦНОљХЈЖШВЂЮоУїЯдБфЛЏЁЃетБэУїУЋЯИбЊЙмЁАЦНОљУмЖШЁБИХФюЕФгІгУЗЖЮЇЪЧгаЯоЕФЃЌКмЖрЪБКђЦфЗжВМзДПіЦ№зХживЊЕФзїгУЁЃМЦЫуНсЙћЛЙБэУїЃЌОрУЋЯИбЊЙмДѓгк100 ІЬmЕФЕиЗНЃЌдкздТЩдЫЖЏЕФећИіжмЦкФкЃЌМИКѕЖМДІгкШБбѕзДЬЌЁЃЮФЯз[20]жИГіЃЌе§ГЃШЫЬхЯИАћгыУЋЯИбЊЙмЕФОрРыаЁгк50 ІЬmЁЃ
ШБбѕЧјЯћУ№КѓЃЌМфжЪЯйпАХЈЖШНЕЕЭЃЌВПЗжУЋЯИбЊЙмЧАРЈдММЁжиаТЙиБеЁЃВЛЩйЪЕбщЙлВтЖМЗЂЯждке§ГЃЩњРэЬѕМўЯТЃЌУЋЯИбЊЙмФкЕФСїЖЏЪБЖЯЪБајЃЌДѓаЁКЭЗНЯђЖМОГЃИФБфЁЃгаЕФбаОПеп[21]ИќЗЂЯжЫљЙлВтЕФШЋВПУЋЯИбЊЙмОљгаГЄЖЬВЛЕШЕФСуСїСПЪБМфЃЌВЂШЯЮЊетЪЧгЩWBCЖТШћдьГЩЕФЁЃЮвУЧЕФбаОПдђБэУїЃЌУЋЯИбЊЙмЕквЛЕїНквВПЩвддьГЩетвЛЯжЯѓЁЃ
, АйФДвНвЉ
ЙњМвздШЛПЦбЇЛљН№зЪжњЯюФП(19472067)
ВЮПМЮФЯз
1ЃЌЮтЦфЯФжїБр.ЬхвКвђЫиКЭбЊвКбЛЗВЁРэЩњРэбЇ.ББОЉЃКББОЉвНПЦДѓбЇЁЂжаЙњаКЭвНПЦДѓбЇСЊКЯГіАцЩчЃЌ1991
2ЃЌGranger HJ,Borders JLЃЌMeininger GA et al.Microcirculatory control systems. in: The physiology and pharmacology of the microcirculation, vol.1, Mortillaro, NA(ed), Academic Press,1983ЁУ209
3ЃЌGranger HJЃЌShepherd AP. Dynamics and control of the microcirculation. Adv.in Biomed. Eng.1979;7ЁУ1
, АйФДвНвЉ
4ЃЌTakahashi EЃЌDoi K. Regulation of oxygen diffusion in hypoxic isolated cardiac myocytes. Am. J. Physiol,1996ЃЛ271ЁУH1734
5ЃЌHoshi Y, Hazeki OЃЛTamura M. Oxygen dependence of redox state of copper in cytochrome oxydase in vitro. J.Appl.Physiol.1993;74ЁУ1622
6ЃЌHester RL. Uptake of metabolites by postcapillary venules: Mechanism for the control of arteriolar diameter. Microvasc. Res,1993;46(2)ЁУ254
7ЃЌDietrick HHЃЌTyml K. Capillary as a communicating medium in the microvasculature. Microvasc. Res.1992ЃЛ43(1)ЁУ87
, АйФДвНвЉ
8ЃЌMitchell DЃЌYu KЃЌTyml K. Comparable effects of arteriolar and capillary stimuli on blood flow in rat skeletal muscle. Microvasc. Res.1997;53(1)ЁУ22
9ЃЌBertuglia S,Colantuoni AЃЌArnold M et al.Dynamic coherence analysis of vasomotion and flow motion in skeletal muscle microcirculation. Microvasc. Res,1996;52(3)ЁУ235
10ЃЌDuran WN,Mortimer PNЃЌLemich JM et al.Capillary reserve in isometrically contracting dog hearts. Am J. Physiol.1977ЃЛ233ЁУH276
, АйФДвНвЉ
11ЃЌAschheim E.Passage of substances across the walls of blood vessels: kinetics and mechanism. in: Microcirculation. vol.1,Kaley, G et al.(eds).Univ.Park Press. Baltimore, London,Tokyo,1977ЁУ213
12ЃЌLee SЃЌSchmid-Schonbein GW. Biomechanical model for the myogenic response in the microcirculation:Part I-Formulation and initial testing. J.Biomech.Eng.1996;118ЁУ145
13ЃЌFrame MDSЃЌSarelius IH. Arteriolar bifurcation angles vary with position and when flow is changed. Microvasc.Res,1993;46(2)ЁУ190
, http://www.100md.com
14ЃЌKontos HA,Fingar MHЃЌWieman JY et al.Role of tissue hypoxia in local regulation of cerebrol microcirculation. Am. J. Physiol,1978;234ЁУH582
15ЃЌKunert MP, Liard JFЃЌAbraham DJ et al.Low-affinity hemogl-
obin increases tissue PO2 and decreases arteriolar diameter and flow in the rat cremaster muscle. Microvasc. Res.1996;52(1)ЁУ58
16ЃЌKroll KЃЌStepp DW. Adenosine kinetics in canine coronary circulation. Am J. Physiol.1996;270(4)ЁУH1469
, http://www.100md.com
17ЃЌBoegehold MA. Enhanced arteriolar vasomotion in rats with chronic salt-induced hypertension. Microvasc.Res.1993;45(1)ЁУ83
18ЃЌDirnagl U, Lindauer UЃЌVillringer A. Nitric oxide synthase blockade enhances vasomotion in the cerebral microcirculation of anesthetized rats. Microvasc.Res.1993;45(3)ЁУ318
19ЃЌЬя ХЃ.ЮЂбЛЗ.ББОЉЃКПЦбЇГіАцЩчЃЌ1986ЁУ1
20ЃЌЮфаЧЛЇБр.ШЫЬхе§ГЃЩњРэЪ§Он.ББОЉЃКПЦбЇЦеМАГіАцЩчЃЌ1987ЁУ55
21ЃЌMayrovitz MN. Skin capillary matrics and hemodynamics in the hairless mouse. Microvasc.Res,1992;43(1)ЁУ46
(ЪеИхЃК1998-12-17), АйФДвНвЉ