人胃癌细胞中COX-2基因的表达及其生物学活性
作者:李玲 吴开春 吴汉平 王新 聂勇战 樊代明
单位:李玲(第四军医大学西京医院全军消化病研究所,陕西西安,710033);吴开春(第四军医大学西京医院全军消化病研究所,陕西西安,710033);吴汉平(第四军医大学西京医院全军消化病研究所,陕西西安,710033);王新(第四军医大学西京医院全军消化病研究所,陕西西安,710033);聂勇战(第四军医大学西京医院全军消化病研究所,陕西西安,710033);樊代明(第四军医大学西京医院全军消化病研究所,陕西西安,710033)
关键词:环氧合酶2;表达;前列腺素E2;肿瘤
细胞与分子免疫学杂志000325
摘要:目的为研究环氧合酶2(cyclooxygenase2,COX-2)或通过前列腺素(prostaglandin,PG)引起的肿瘤免疫机制,我们检测了人胃癌细胞中COX-2mRNA及蛋白质水平的表达,PGE2的释放和COX-2酶活性的抑制剂消炎痛(indomethacin,Indo)对PGE2的释放和肿瘤细胞生长的影响。方法采用免疫细胞化学染色、RTPCR,ELISA,以及MTT比色试验方法研究人胃癌细胞COX-2基因与蛋白质的表达、PGE2释放以及消炎痛对PGE2释放和细胞活力的影响。结果所有检测的胃癌细胞均表达COX-2mRNA,胃癌细胞MKN45,SGC7901和AGS中有COX-2蛋白质的表达,MGC803则不表达。ELISA表明,所有胃癌细胞均有PGE2释放,给予COX-2表达的诱导剂PMA后,MGC803中PGE2的释放由57μg/L增加至63μg/L,小剂量COX-2酶活性的抑制剂—消炎痛(0.125μmol/L)则可使其水平降至47μg/L。MTT法进一步显示,消炎痛可使肿瘤细胞增殖与活力明显抑制,尤以小剂量为著。结论人胃癌细胞中存在COX-2基因的表达、PGE2的释放及肿瘤细胞增殖之间的相互关联。
, http://www.100md.com
中图号:R735.2 文献标识码:A 文章编号:1007-8738(2000)03-0255-05
Expression and biological activity of COX-2 gene in human
gastric cancer cell lines
LI Ling WU Kai-chun WU Han-ping WANG Xin NIE Yong-zhan FAN Dai-ming
(Institute of Digestive Diseases, Xijing Hosiptal,Fourth Military Medical University,Xi'an 710033,Shaanxi Province,China)
, 百拇医药
Abstract: Aim To detect the expression of COX-2, the level of PGE2 in human gastric cancer cell lines and the inhibition of indomethacin on cell growth and PGE2 level, so to elucidate the role of cyclooxygenase 2 (COX-2) and prostaglandin (PG) in tumor immunity. Methods Immunohistochemistry, RT PCR, MTT assay and ELISA were used. Results COX-2 mRNA expression was observed in all gastric cancer cell lines (MKN45, SGC7901, AGS, MGC803);but COX-2 protein expression was only shown in MKN45, SGC7901 and AGS. ELISA exhibited that all gastric cancer cell lines produced PGE2, and PMA increased the level of PGE2 in MGC803 (from 57 μ g/L to 63 μ g/L), while 0.125 μ mol/L indomethacin decreased the level to 47 μ g/L. Indomethacin inhibited proliferation and activity of SGC7901 in MTT assay. Conclusion COX-2 gene expression exists in human gastric cancer cell lines, and there seems to be correlation between COX-2 expression, PGE2 production and cell proliferation.
, http://www.100md.com
Keywords: COX-2; expression; PGE2; tumor■
环氧合酶-2(cyclooxygenase2,COX-2)是前列腺素(prostaglandin,PG)生物合成过程中的一个重要限速酶,可将花生四烯酸代谢为各种PG产物,从而维持机体的病理生理过程。生理状态下PG由单核-巨噬细胞系统产生,以调节细胞因子的平衡。炎症时诱导IL-6产生的物质同时可诱导COX-2mRNA的产生,而且在炎症部位IL-6和COX-2mRNA表达同时上调,从而调动机体对炎症的免疫应答;而应用COX-2的选择性抑制剂(NS-398)则可使IL-6的产生受抑制〔1〕。已有大量研究报道,肿瘤组织(如头颈、胸、肺和结肠等)可产生较高水平的PG。吴开春等〔2〕曾报道,同正常组织相比较,胃癌组织产生PGE2的水平增高。PG在肿瘤发生中的作用表现为:影响有丝分裂,促进细胞粘附,抑制凋亡发生及减低免疫监视机能〔3〕。肿瘤组织PG产生的增加常伴有COX活性,尤其是COX-2活性的增加,同时可有COX-2表达增加。我们应用免疫组化染色显示,胃癌组织COX-2蛋白的表达(65.5%)显著高于浅表性胃炎(18.2%)。为进一步了解基因水平上COX-2的表达,探讨COX2,PG与肿瘤这三者之间的关系,我们采用免疫组化染色,RT-PCR,ELISA和MTT等方法,检测了COX2的表达和PGE2的释放,并分析了COX-2表达的诱导剂及酶活性的抑制剂对PGE2释放和细胞增殖的影响。
, 百拇医药
1 材料和方法
1.1材料人胃癌细胞MKN45,SGC7901,AGS和MGC803均为本所保存。RPMI1640,DMEM
培养基及DMEM/F12培养基购自Gibco公司。胎牛血清购自Hyclone公司。兔抗人COX1和COX2多克隆抗体购自Cayman公司。SABC免疫酶染色试剂盒及DAB显色试剂盒,购自武汉博士德公司。总RNA超绝UNIQ10抽提试剂盒及Taq酶,购自NSBCSangon公司。MMuLv购自MBI公司。100bpDNAladder购自华美生物工程公司。PGE2ELISA试剂盒购自Amersham公司。PMA及消炎痛为Sigma公司产品。
1.2方法
1.2.1RTPCR①细胞的收集:取对数生长期的细胞(有血清或无血清),用血细胞计数器计数达2×107时,用2.5g/L胰蛋白酶消化,制成单细胞悬液,经500r/min离心10min,收集细胞沉淀,再以0.01mmol/LpH7.4PBS洗涤沉淀1次,于70℃保存。②细胞总RNA的提取:应用NSBC超绝UNIQ10柱总RNA抽提试剂盒,将抽提的RNA样品进行甲醛变性胶电泳了解其完整性,并用分光光度计测定A260nm及A280nm值分析其纯度,同时进行定量。③cDNA的合成:于冰上混合1μgRNA样品及1.2μL随机六聚体引物,于70℃退火5min后,立即置于冰浴中,加入4μL逆转录酶缓冲液,1μL20~40Mu/LRNasin及2μL10mmol/LdNTP,充分混合,25℃5min;最后加入2μLMMuLV逆转录酶,补加水至20μL,于25℃5min,37℃60min,70℃加热10min终止反应,4℃保存。④PCR:引物序列hCOX2为:
, http://www.100md.com
P1:5′TAGAACCCACTCCAAACACAG3′;
P2:5′TCATCAGGCACAGGAGGAAG3′.
PCR扩增片段长度为232bp;反应体系为:0.8μLcDNA,2μL10×Taq酶缓冲液,0.4μL10mmol/LdNTP,0.8μL25mmol/LMgCl2,上下游引物各0.4μL及0.5μLTaq酶,补加水至20μL。分别以去离子水代替模板作为阴性对照,以人结肠癌细胞Caco2之模板为阳性对照。PCR条件:95℃变性45s,54℃复性90s,72℃延伸90s,共30个循环。最后于54℃延伸2min,72℃延伸3min。反应完毕后,取10μL扩增产物,于17g/L的琼脂糖凝胶上进行电泳分析。为比较各细胞间表达水平的强弱,应用图象分析仪对扩增条带做了扫描半定量。
1.2.2免疫细胞化学染色①细胞的培养与固定:人胃腺癌细胞系(MKN45,SGC7901,AGS和MGC803)的培养液,为RPMI1640(含100mL/L热灭活胎牛血清)或为DMEM/F12无血清培养基,于37℃,50mL/LCO2孵箱中培养。待细胞生长至70%~80%时,用2.5g/L胰蛋白酶和0.2g/LEDTA消化,制备单细胞悬液,接种于载玻片上,置37℃培养8~10h。待细胞贴壁后,取出,用冷丙酮于室温固定10min,20℃保存。②SABC法染色:主要步骤为:3mL/LH2O2及甲醇室温30min,3g/LTritonX10037℃12min,30mL/L正常羊血清室温封闭2h。依次加1:125的兔抗人COX1和2多抗(置湿盒内4℃过夜),1:100生物素化的羊抗兔IgG(37℃1h)及ABC复合物(37℃40min),最后以DAB显色并封片、镜检。实验中以同等稀释度的兔血清及PBS代替一抗作为阴性对照,以人结肠癌细胞Caco2作为阳性标本对照。
, http://www.100md.com
1.2.3ELISA以SGC7901和MGS803为研究对象,分为a,b,c3个组,分别接种于9cm的培养皿。a组加同等稀释度的DMSO,b组加25μg/LPMA孵育3h,c组加0.125μmol/L消炎痛,孵育24h,至相应的时间段分别收集上清及细胞沉淀。取0.5mL培养上清,加入0.5mL800mL/L乙醇及10μL冰醋酸轻微混合,室温静置5min,以2500r/min离心2min,收集上清。取105细胞沉淀,用PBS洗涤两次,加入100μL细胞裂解液,置室温孵育10min,用加样器反复抽吸混匀。所制备的样品均须立即使用。将50μL标准品和等量样品分别加至相应孔中,再加入50μL稀释的PGE2抗血清和50μL稀释的PGE2结合物,于20℃孵育1h。取出,用洗液洗涤4次,立即加入150μLTMB底物,25℃孵育30min,用100μL1mol/L硫酸终止反应,在酶联免疫检测分析仪读取A450nm的值。每一标准品及样品均设置复孔,不加样品的孔用于调零。以检测缓冲液(培养上清)或裂解缓冲液(细胞裂解液)作为非特异性对照,以%B/B0(y轴)对PGE2浓度的对数值(x轴)作标准曲线,计算相关系数(r)为99%,求解回归方程,得出各样品中PGE2浓度的对数值。%B/B0的计算公式为:
, 百拇医药
标准品或样品的A值—非特异性对照的A值
%B/B0=()×100%
零标准品的A值—非特异性对照的A值
1.2.4MTT比色试验用2.5g/L胰蛋白酶消化单层培养细胞SGC7901,制备单细胞悬液。以每孔3×103个细胞接种于96孔培养板(每孔200μL),分别加消炎痛(2,1,0.5,0.25及0.125μmol/L)或不加消炎痛,移入37℃50mL/LCO2孵箱中培养24h。然后每孔加入MTT溶液(5g/L)20μL,于37℃继续孵育4h,终止培养。吸弃孔中的培养上清,加入150μLDMSO振荡10min,在酶联免疫检测分析仪测定A490nm的值。以只加培养液不加细胞的空白对照孔调零。
2 结果
2.1人胃癌细胞COX2mRNA的表达四株胃癌细胞的RTPCR产物经电泳分析,可见1条约232bp的扩增带,特异性好,阴性对照无相应条带出现(图1)。采用无血清培养基DMEM/F12培养后,仍有232bp的特异性扩增条带,只是条带的亮度稍下降(1.7与1.59)(图2),说明胃癌细胞中有COX2基因的表达。同时,检测的另外4株细胞系亦有COX2基因的表达(图1),但与正常肝细胞相比较,其表达水平稍弱。
, 百拇医药
图1 COX-2mRNA在肿瘤细胞中的表达
图2 FCS对COX-2mRNA表达的影响
2.2人胃癌细胞COX-2蛋白的表达SABC检测结果显示,COX-2和1在胃癌细胞系MKN45,SGC7901和AGS中的表达呈阳性,MGC803不表达COX-2,对COX-1的表达呈弱阳性。阳性物质呈棕黄色颗粒状,主要弥漫分布于整个胞浆或沿核膜周边呈线状分布(图3)。两种COX表达的强度相比较,COX-2的表达明显高于COX-1。采用无血清培养基DMEM/F12培养后,血清对于COX-2表达的干扰因素去除,仍可见有COX-2的表达。作为对照的结肠癌细胞Caco-2表达COX-2较高;而肝癌细胞HCC7721和白血病细胞K562则不表达COX-2和COX-1。
, 百拇医药
图3COX-2的免疫组化染色
2.3PGE2的释放所有的胃癌细胞如MKN45,AGS,SGC7901和MGC803均有PGE2的释放,但释放的量差异不显著,培养上清中PGE2的水平波动
在10~20μg/L之间。对不表达COX2的MGC803细胞而言,COX2表达的诱导剂(25μg/L的PMA)可使细胞内PGE2的水平由59μg/L增加至63μg/L,小剂量(0.125μmol/L)的消炎痛则可使细胞内PGE2的水平下降到47μg/L(图4)。培养上清亦有相同的趋势。PMA对SGC7901释放PGE2的作用微弱,甚至表现为抑制效应。直接用0.125μmol/L的消炎痛亦可抑制PGE2的释放,但其抑制效应没有MGC803明显(图5)。无论何种处理,由细胞内释放出的PGE2水平要明显高于培养上清。但就MGC803而言,这种差异明显(由11.9μg/L至59.4μg/L);而SGC7901则在19.31~33.36μg/L之间。
, 百拇医药
图4COX-2诱导剂与抑制剂对MGC803细胞内及培养上清中PGE2产生的影响
图5消炎痛对SGC7901细胞内及培养上清中PGE2释放的影响
2.4MTT比色试验不同浓度的消炎痛其在A490nm的值与对照组SGC7901相比较,均有不同程度的下降(介于2~0.25μmol/L浓度之间),A490nm值相差不显著;而0.125μmol/L的消炎痛则可明显降低A490nm值,使细胞活力与增殖明显抑制(图6)。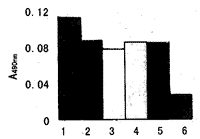
图6不同浓度的消炎痛对SGC7901细胞增殖与活性的影响
3 讨论
, 百拇医药
我们发现,人胃癌细胞中有mRNA和mCOX-2蛋白的表达,去除血清干扰后COX-2表达仍存在,提示胃癌细胞中有COX-2基因的过表达。这一结果与文献报道相一致,即COX-2mRNA及其蛋白质水平与正常组织相比较显著升高,而COX-1水平则无明显改变。如在结肠直肠腺瘤及腺癌〔4,5〕、胃癌〔6〕、胰腺癌〔7〕、肺癌〔8〕、前列腺癌〔9〕和骨肉瘤〔10〕中,COX2的表达升高。我们检测的另1株人白血病细胞K562中,同样有COX2mRNA的过表达。此外,非甾体抗炎药(NSAID)可抑制人与实验动物结肠、食管、胰腺、乳腺、肺、皮肤、膀胱和子宫颈肿瘤的发生,因此推测,COX-2表达的上调可能与肿瘤的发生有关。我们还发现,人正常肝细胞(L02)中,COX-2mRNA的表达水平高于人肝癌细胞系HCC7721。虽然就HCC7721细胞系而言,COX-2的表达较COX-1的表达上调,这似乎与对胃癌的研究结果相矛盾,可能其在人正常肝细胞为组成性表达,也可能肝癌细胞对COX-2的表达水平同正常比较并无升高。也就是说,在肝癌的发生中,无COX-2的参与。但无论如何,COX-2表达的上调是一种普遍现象,而其是否参与所有肿瘤的发生,尚难下结论。可以肯定的是,在胃癌的发生中有COX-2的参与。
, 百拇医药
ELISA分析胃癌细胞中有PGE2的释放,提示胃癌细胞有COX的环氧化酶活性和过氧化物酶活性,其来源可能为COX-2,也可能为COX-1。应用PMA及小剂量的消炎痛诱导COX-2的表达或抑制COX-2的酶活性,MGC803对PGE2的释放亦相应增加或减少,同时表达COX-2水平较高的SGC7901对PGE2的释放也可被小剂量消炎痛抑制,表明在胃癌细胞,尤其是高表达COX-2的胃癌细胞PGE2的产生源自COX-2。MTT检测表明,消炎痛可降低SGC7901细胞的A490nm值,抑制细胞的增殖与活性,可能是其抑制了COX的表达与酶活性所致。小剂量的消炎痛作用更显著,表明其抑制了两种异构酶中的COX-2,因为小剂量消炎痛主要抑制COX-2。由此可见,COX-2的表达,PGE2的释放和肿瘤细胞的增殖之间有一定的相关性。因此,如选用高选择性COX-2的抑制剂,将对PGE2的释放和肿瘤细胞的增殖产生显著的作用,即对于胃癌的治疗具有潜在的意义。
PMA可通过蛋白激酶C途径而诱导COX-2的表达,使COX2酶活性增加,进一步导致细胞增殖及PGE2释放,其对不表达COX-2的MGC803细胞的效应显著;而对本身可表达COX-2的SGC7901,其作用很微弱甚至表现为抑制。有可能在SGC7901中,COX-2的转录已启动或已处于转录激活状态,此时PMA对其影响的作用很弱。MGC803细胞内与培养上清中PGE2水平的差异悬殊,而SGC7901则不显著,可能为同样的原因,即由于SGC7901中COX-2的转录已经启动,其酶活性催化的PGE2已多数释放至上清中。MGC803不表达COX--2,释放至上清中的PGE2水平则低。这种低水平的PGE2可能为PG产生过程中的另一限速酶COX-1引起。
, http://www.100md.com
有关COX-2对肿瘤免疫的影响,文献报道不多。Urata等〔11〕发现,人胃癌细胞COX-2呈过表达时,可增强淋巴细胞浸润与转移。Tsujii发现〔12〕,结肠癌细胞表达的COX-2可抑制中性粒细胞浸润和巨噬细胞活化,从而导致抗肿瘤免疫监视机能抑制。另外,在非小细胞肺癌中,COX-2阳性的肿瘤细胞可引起细胞因子失衡:即增加巨噬细胞、淋巴细胞产生IL-10,抑制巨噬细胞产生IL-12。IL-12可诱导Th1细胞产生Th1型细胞因子,介导有效的抗肿瘤免疫应答〔8〕。COX-2也可通过PG在肿瘤免疫中发挥其作用:如PGE2可抑制具有免疫调节功能的淋巴因子产生,抑制T细胞和B细胞增殖,T细胞母细胞化及抑制NK细胞、LAK细胞和CTL的细胞毒活性。同时,PGE2还可抑制TNF的产生,诱导IL-10产生。因此,应用环氧化酶抑制剂(如阿斯匹林、苏灵达和消炎痛等)可减低肿瘤引起的免疫抑制作用〔2〕。■
Fig 1 Expression of COX 2 mRNA in tumor cells 1: Negative control;2:fbs; 3:HCC7721; 4:K562; 5:Lo2;6:MGC803; 7:AGS; 8:SGC7901; 9:MKN45;10:100 bp DNA ladder; 11:Caco 2.
, 百拇医药
Fig 2 FCS′ s influence on COX 2 mRNA expression 1: Negative control; 2:SGC7901 10% FCS; 3:AGS 10% FCS; 4:100 bp DNA ladder; 5:AGS DMEM/F12; 6:SGC7901 DMEM/F12.
Fig 3 Immunohistochemical staining of COX 2 (× 40) A: COX 1; B: COX 2; C: Rabbit IgG.
Fig 4 The influence of COX 2 inductor and inhibitor on produ ction of PGE2 from MGC803 cells and in culture supernatant
Fig 5 The influence of indomethacin on release of PGE2 from SGC7901 cells and in culture supernatant
, http://www.100md.com
Fig 6 The influence of indomethacin of different concentrations on SGC7901 cell proliferation and activity 1~ 6: Concentrations of indomathacin are successively 0,2,1,0.5,0.25 and 0.125 μ mol/L.
基金项目:国家自然科学基金资助,No.39870230
作者简介:李玲,女,27岁,硕士生西安市长乐西路15号,Tel.(029)3375229 2000 by J Cell Mol Immunol Press www.immunology. net/jcmi; Email. immuedit@fmmu.edu.cn
参考文献:
, http://www.100md.com 〔1〕 Hinson R, Williams JA, Shacter E. Elevated interleukin 6 is induced by prostaglandin E2 in a murine model of inflammation: possible role of cyclooxygenase 2〔J〕 . Proc Natl Acad Sci USA, 1996; 93(10):4885- 4890.
〔2〕 吴 开 春 , 邓 敬 兰 , 张 学 庸 .胃 癌 组 织 PGE产 生 及 其 对 PHA介 导 的 3H TdR掺 入 的 抑 制 〔J〕 .第 四 军 医 大 学 学 报 , 1988; 9 (5): 295- 296.
〔3〕 Subbaramiah K, Zakim D, Weksler BB, et al. Inhibition of cyclooxygenase: a novel approach to cancer prevention〔J〕 . Proc Soc Exp Biol Med, 1997; 216 (2):201- 210.
, http://www.100md.com
〔4〕 Eberhart CE, Coffey RJ, Radhika A, et al. Up regulation of cyclooxygenase 2 gene expression in human colorectal adenomas and adenocarcinomas〔J〕 . Gastroenterology, 1994; 107(4):1183- 1188.
〔5〕 H, Kawahito Y, Wilder RL, et al. Expression of cyclooxygenase 1 and 2 in human colorectal cancer〔J〕 . Cancer Res, 1995; 55 (17):3785- 3789.
〔6〕 Ristimaki A, Honkanen N, Jankala H, et al. Expression of cyclooxygenase 2 in human gastric carcinoma〔J〕 . Cancer Res, 1997; 57 (7):1276- 1280.
, 百拇医药
〔7〕 Tucker ON, Dannenberg AJ, Yang EK, et al. Cyclooxygenase 2 expression is up regulated in human pancreatic cancer〔J〕 . Cancer Res, 1999; 59 (5): 987- 990.
〔8〕 Huang M, Stolina M, Sharma S, et al. Non small cell lung cancer cyclooxygenase 2 dependent regulation of cytokine balance in lymphocytes and macrophages: up regulation of interleukin 10 and down regulation of interleukin 12 production〔J〕 . Cancer Res, 1998; 58 (7):1208- 1216.
, 百拇医药
〔9〕 Tjandrawinata RR, Dahiya R, Hughes Fulford M. Induction of cyclooxygenase 2 mRNA by prostaglandin E2 in human prostatic carcinoma cells〔J〕 . Br J Cancer, 1997; 75 (8):1111- 1118.
〔10〕 Wong E, Deluca C, Boily C, et al. Characterization of autocrine inducible prostaglandin H synthase 2 (PGHS 2) in human osteos arcoma cells〔J〕 . Inflamm Res, 1997; 46 (1): 51- 56.
〔11〕 Urata H, Kawano S, Tsujii S, et al. Enhanced lymphatic invasion and metastasis with cyclooxygenase 2 overexpression in human gastric carcinoma〔J〕 . Gastroenterology, 1998; 114(4pt2): A798.
〔12〕 Tsujii M, Kawano S, Tsujii S, et al. Effect of cyclooxygenase 2 expression in colon cancer cells on chemotaxis of neutrophils and macrophage〔J〕 . Gastroenterology, 1998; 114(4pt2): A797.
收稿日期:2000-01-02
修回日期:2000-02-16, 百拇医药
单位:李玲(第四军医大学西京医院全军消化病研究所,陕西西安,710033);吴开春(第四军医大学西京医院全军消化病研究所,陕西西安,710033);吴汉平(第四军医大学西京医院全军消化病研究所,陕西西安,710033);王新(第四军医大学西京医院全军消化病研究所,陕西西安,710033);聂勇战(第四军医大学西京医院全军消化病研究所,陕西西安,710033);樊代明(第四军医大学西京医院全军消化病研究所,陕西西安,710033)
关键词:环氧合酶2;表达;前列腺素E2;肿瘤
细胞与分子免疫学杂志000325
摘要:目的为研究环氧合酶2(cyclooxygenase2,COX-2)或通过前列腺素(prostaglandin,PG)引起的肿瘤免疫机制,我们检测了人胃癌细胞中COX-2mRNA及蛋白质水平的表达,PGE2的释放和COX-2酶活性的抑制剂消炎痛(indomethacin,Indo)对PGE2的释放和肿瘤细胞生长的影响。方法采用免疫细胞化学染色、RTPCR,ELISA,以及MTT比色试验方法研究人胃癌细胞COX-2基因与蛋白质的表达、PGE2释放以及消炎痛对PGE2释放和细胞活力的影响。结果所有检测的胃癌细胞均表达COX-2mRNA,胃癌细胞MKN45,SGC7901和AGS中有COX-2蛋白质的表达,MGC803则不表达。ELISA表明,所有胃癌细胞均有PGE2释放,给予COX-2表达的诱导剂PMA后,MGC803中PGE2的释放由57μg/L增加至63μg/L,小剂量COX-2酶活性的抑制剂—消炎痛(0.125μmol/L)则可使其水平降至47μg/L。MTT法进一步显示,消炎痛可使肿瘤细胞增殖与活力明显抑制,尤以小剂量为著。结论人胃癌细胞中存在COX-2基因的表达、PGE2的释放及肿瘤细胞增殖之间的相互关联。
, http://www.100md.com
中图号:R735.2 文献标识码:A 文章编号:1007-8738(2000)03-0255-05
Expression and biological activity of COX-2 gene in human
gastric cancer cell lines
LI Ling WU Kai-chun WU Han-ping WANG Xin NIE Yong-zhan FAN Dai-ming
(Institute of Digestive Diseases, Xijing Hosiptal,Fourth Military Medical University,Xi'an 710033,Shaanxi Province,China)
, 百拇医药
Abstract: Aim To detect the expression of COX-2, the level of PGE2 in human gastric cancer cell lines and the inhibition of indomethacin on cell growth and PGE2 level, so to elucidate the role of cyclooxygenase 2 (COX-2) and prostaglandin (PG) in tumor immunity. Methods Immunohistochemistry, RT PCR, MTT assay and ELISA were used. Results COX-2 mRNA expression was observed in all gastric cancer cell lines (MKN45, SGC7901, AGS, MGC803);but COX-2 protein expression was only shown in MKN45, SGC7901 and AGS. ELISA exhibited that all gastric cancer cell lines produced PGE2, and PMA increased the level of PGE2 in MGC803 (from 57 μ g/L to 63 μ g/L), while 0.125 μ mol/L indomethacin decreased the level to 47 μ g/L. Indomethacin inhibited proliferation and activity of SGC7901 in MTT assay. Conclusion COX-2 gene expression exists in human gastric cancer cell lines, and there seems to be correlation between COX-2 expression, PGE2 production and cell proliferation.
, http://www.100md.com
Keywords: COX-2; expression; PGE2; tumor■
环氧合酶-2(cyclooxygenase2,COX-2)是前列腺素(prostaglandin,PG)生物合成过程中的一个重要限速酶,可将花生四烯酸代谢为各种PG产物,从而维持机体的病理生理过程。生理状态下PG由单核-巨噬细胞系统产生,以调节细胞因子的平衡。炎症时诱导IL-6产生的物质同时可诱导COX-2mRNA的产生,而且在炎症部位IL-6和COX-2mRNA表达同时上调,从而调动机体对炎症的免疫应答;而应用COX-2的选择性抑制剂(NS-398)则可使IL-6的产生受抑制〔1〕。已有大量研究报道,肿瘤组织(如头颈、胸、肺和结肠等)可产生较高水平的PG。吴开春等〔2〕曾报道,同正常组织相比较,胃癌组织产生PGE2的水平增高。PG在肿瘤发生中的作用表现为:影响有丝分裂,促进细胞粘附,抑制凋亡发生及减低免疫监视机能〔3〕。肿瘤组织PG产生的增加常伴有COX活性,尤其是COX-2活性的增加,同时可有COX-2表达增加。我们应用免疫组化染色显示,胃癌组织COX-2蛋白的表达(65.5%)显著高于浅表性胃炎(18.2%)。为进一步了解基因水平上COX-2的表达,探讨COX2,PG与肿瘤这三者之间的关系,我们采用免疫组化染色,RT-PCR,ELISA和MTT等方法,检测了COX2的表达和PGE2的释放,并分析了COX-2表达的诱导剂及酶活性的抑制剂对PGE2释放和细胞增殖的影响。
, 百拇医药
1 材料和方法
1.1材料人胃癌细胞MKN45,SGC7901,AGS和MGC803均为本所保存。RPMI1640,DMEM
培养基及DMEM/F12培养基购自Gibco公司。胎牛血清购自Hyclone公司。兔抗人COX1和COX2多克隆抗体购自Cayman公司。SABC免疫酶染色试剂盒及DAB显色试剂盒,购自武汉博士德公司。总RNA超绝UNIQ10抽提试剂盒及Taq酶,购自NSBCSangon公司。MMuLv购自MBI公司。100bpDNAladder购自华美生物工程公司。PGE2ELISA试剂盒购自Amersham公司。PMA及消炎痛为Sigma公司产品。
1.2方法
1.2.1RTPCR①细胞的收集:取对数生长期的细胞(有血清或无血清),用血细胞计数器计数达2×107时,用2.5g/L胰蛋白酶消化,制成单细胞悬液,经500r/min离心10min,收集细胞沉淀,再以0.01mmol/LpH7.4PBS洗涤沉淀1次,于70℃保存。②细胞总RNA的提取:应用NSBC超绝UNIQ10柱总RNA抽提试剂盒,将抽提的RNA样品进行甲醛变性胶电泳了解其完整性,并用分光光度计测定A260nm及A280nm值分析其纯度,同时进行定量。③cDNA的合成:于冰上混合1μgRNA样品及1.2μL随机六聚体引物,于70℃退火5min后,立即置于冰浴中,加入4μL逆转录酶缓冲液,1μL20~40Mu/LRNasin及2μL10mmol/LdNTP,充分混合,25℃5min;最后加入2μLMMuLV逆转录酶,补加水至20μL,于25℃5min,37℃60min,70℃加热10min终止反应,4℃保存。④PCR:引物序列hCOX2为:
, http://www.100md.com
P1:5′TAGAACCCACTCCAAACACAG3′;
P2:5′TCATCAGGCACAGGAGGAAG3′.
PCR扩增片段长度为232bp;反应体系为:0.8μLcDNA,2μL10×Taq酶缓冲液,0.4μL10mmol/LdNTP,0.8μL25mmol/LMgCl2,上下游引物各0.4μL及0.5μLTaq酶,补加水至20μL。分别以去离子水代替模板作为阴性对照,以人结肠癌细胞Caco2之模板为阳性对照。PCR条件:95℃变性45s,54℃复性90s,72℃延伸90s,共30个循环。最后于54℃延伸2min,72℃延伸3min。反应完毕后,取10μL扩增产物,于17g/L的琼脂糖凝胶上进行电泳分析。为比较各细胞间表达水平的强弱,应用图象分析仪对扩增条带做了扫描半定量。
1.2.2免疫细胞化学染色①细胞的培养与固定:人胃腺癌细胞系(MKN45,SGC7901,AGS和MGC803)的培养液,为RPMI1640(含100mL/L热灭活胎牛血清)或为DMEM/F12无血清培养基,于37℃,50mL/LCO2孵箱中培养。待细胞生长至70%~80%时,用2.5g/L胰蛋白酶和0.2g/LEDTA消化,制备单细胞悬液,接种于载玻片上,置37℃培养8~10h。待细胞贴壁后,取出,用冷丙酮于室温固定10min,20℃保存。②SABC法染色:主要步骤为:3mL/LH2O2及甲醇室温30min,3g/LTritonX10037℃12min,30mL/L正常羊血清室温封闭2h。依次加1:125的兔抗人COX1和2多抗(置湿盒内4℃过夜),1:100生物素化的羊抗兔IgG(37℃1h)及ABC复合物(37℃40min),最后以DAB显色并封片、镜检。实验中以同等稀释度的兔血清及PBS代替一抗作为阴性对照,以人结肠癌细胞Caco2作为阳性标本对照。
, http://www.100md.com
1.2.3ELISA以SGC7901和MGS803为研究对象,分为a,b,c3个组,分别接种于9cm的培养皿。a组加同等稀释度的DMSO,b组加25μg/LPMA孵育3h,c组加0.125μmol/L消炎痛,孵育24h,至相应的时间段分别收集上清及细胞沉淀。取0.5mL培养上清,加入0.5mL800mL/L乙醇及10μL冰醋酸轻微混合,室温静置5min,以2500r/min离心2min,收集上清。取105细胞沉淀,用PBS洗涤两次,加入100μL细胞裂解液,置室温孵育10min,用加样器反复抽吸混匀。所制备的样品均须立即使用。将50μL标准品和等量样品分别加至相应孔中,再加入50μL稀释的PGE2抗血清和50μL稀释的PGE2结合物,于20℃孵育1h。取出,用洗液洗涤4次,立即加入150μLTMB底物,25℃孵育30min,用100μL1mol/L硫酸终止反应,在酶联免疫检测分析仪读取A450nm的值。每一标准品及样品均设置复孔,不加样品的孔用于调零。以检测缓冲液(培养上清)或裂解缓冲液(细胞裂解液)作为非特异性对照,以%B/B0(y轴)对PGE2浓度的对数值(x轴)作标准曲线,计算相关系数(r)为99%,求解回归方程,得出各样品中PGE2浓度的对数值。%B/B0的计算公式为:
, 百拇医药
标准品或样品的A值—非特异性对照的A值
%B/B0=()×100%
零标准品的A值—非特异性对照的A值
1.2.4MTT比色试验用2.5g/L胰蛋白酶消化单层培养细胞SGC7901,制备单细胞悬液。以每孔3×103个细胞接种于96孔培养板(每孔200μL),分别加消炎痛(2,1,0.5,0.25及0.125μmol/L)或不加消炎痛,移入37℃50mL/LCO2孵箱中培养24h。然后每孔加入MTT溶液(5g/L)20μL,于37℃继续孵育4h,终止培养。吸弃孔中的培养上清,加入150μLDMSO振荡10min,在酶联免疫检测分析仪测定A490nm的值。以只加培养液不加细胞的空白对照孔调零。
2 结果
2.1人胃癌细胞COX2mRNA的表达四株胃癌细胞的RTPCR产物经电泳分析,可见1条约232bp的扩增带,特异性好,阴性对照无相应条带出现(图1)。采用无血清培养基DMEM/F12培养后,仍有232bp的特异性扩增条带,只是条带的亮度稍下降(1.7与1.59)(图2),说明胃癌细胞中有COX2基因的表达。同时,检测的另外4株细胞系亦有COX2基因的表达(图1),但与正常肝细胞相比较,其表达水平稍弱。
, 百拇医药
图1 COX-2mRNA在肿瘤细胞中的表达
图2 FCS对COX-2mRNA表达的影响
2.2人胃癌细胞COX-2蛋白的表达SABC检测结果显示,COX-2和1在胃癌细胞系MKN45,SGC7901和AGS中的表达呈阳性,MGC803不表达COX-2,对COX-1的表达呈弱阳性。阳性物质呈棕黄色颗粒状,主要弥漫分布于整个胞浆或沿核膜周边呈线状分布(图3)。两种COX表达的强度相比较,COX-2的表达明显高于COX-1。采用无血清培养基DMEM/F12培养后,血清对于COX-2表达的干扰因素去除,仍可见有COX-2的表达。作为对照的结肠癌细胞Caco-2表达COX-2较高;而肝癌细胞HCC7721和白血病细胞K562则不表达COX-2和COX-1。
, 百拇医药
图3COX-2的免疫组化染色
2.3PGE2的释放所有的胃癌细胞如MKN45,AGS,SGC7901和MGC803均有PGE2的释放,但释放的量差异不显著,培养上清中PGE2的水平波动
在10~20μg/L之间。对不表达COX2的MGC803细胞而言,COX2表达的诱导剂(25μg/L的PMA)可使细胞内PGE2的水平由59μg/L增加至63μg/L,小剂量(0.125μmol/L)的消炎痛则可使细胞内PGE2的水平下降到47μg/L(图4)。培养上清亦有相同的趋势。PMA对SGC7901释放PGE2的作用微弱,甚至表现为抑制效应。直接用0.125μmol/L的消炎痛亦可抑制PGE2的释放,但其抑制效应没有MGC803明显(图5)。无论何种处理,由细胞内释放出的PGE2水平要明显高于培养上清。但就MGC803而言,这种差异明显(由11.9μg/L至59.4μg/L);而SGC7901则在19.31~33.36μg/L之间。
, 百拇医药
图4COX-2诱导剂与抑制剂对MGC803细胞内及培养上清中PGE2产生的影响
图5消炎痛对SGC7901细胞内及培养上清中PGE2释放的影响
2.4MTT比色试验不同浓度的消炎痛其在A490nm的值与对照组SGC7901相比较,均有不同程度的下降(介于2~0.25μmol/L浓度之间),A490nm值相差不显著;而0.125μmol/L的消炎痛则可明显降低A490nm值,使细胞活力与增殖明显抑制(图6)。
图6不同浓度的消炎痛对SGC7901细胞增殖与活性的影响
3 讨论
, 百拇医药
我们发现,人胃癌细胞中有mRNA和mCOX-2蛋白的表达,去除血清干扰后COX-2表达仍存在,提示胃癌细胞中有COX-2基因的过表达。这一结果与文献报道相一致,即COX-2mRNA及其蛋白质水平与正常组织相比较显著升高,而COX-1水平则无明显改变。如在结肠直肠腺瘤及腺癌〔4,5〕、胃癌〔6〕、胰腺癌〔7〕、肺癌〔8〕、前列腺癌〔9〕和骨肉瘤〔10〕中,COX2的表达升高。我们检测的另1株人白血病细胞K562中,同样有COX2mRNA的过表达。此外,非甾体抗炎药(NSAID)可抑制人与实验动物结肠、食管、胰腺、乳腺、肺、皮肤、膀胱和子宫颈肿瘤的发生,因此推测,COX-2表达的上调可能与肿瘤的发生有关。我们还发现,人正常肝细胞(L02)中,COX-2mRNA的表达水平高于人肝癌细胞系HCC7721。虽然就HCC7721细胞系而言,COX-2的表达较COX-1的表达上调,这似乎与对胃癌的研究结果相矛盾,可能其在人正常肝细胞为组成性表达,也可能肝癌细胞对COX-2的表达水平同正常比较并无升高。也就是说,在肝癌的发生中,无COX-2的参与。但无论如何,COX-2表达的上调是一种普遍现象,而其是否参与所有肿瘤的发生,尚难下结论。可以肯定的是,在胃癌的发生中有COX-2的参与。
, 百拇医药
ELISA分析胃癌细胞中有PGE2的释放,提示胃癌细胞有COX的环氧化酶活性和过氧化物酶活性,其来源可能为COX-2,也可能为COX-1。应用PMA及小剂量的消炎痛诱导COX-2的表达或抑制COX-2的酶活性,MGC803对PGE2的释放亦相应增加或减少,同时表达COX-2水平较高的SGC7901对PGE2的释放也可被小剂量消炎痛抑制,表明在胃癌细胞,尤其是高表达COX-2的胃癌细胞PGE2的产生源自COX-2。MTT检测表明,消炎痛可降低SGC7901细胞的A490nm值,抑制细胞的增殖与活性,可能是其抑制了COX的表达与酶活性所致。小剂量的消炎痛作用更显著,表明其抑制了两种异构酶中的COX-2,因为小剂量消炎痛主要抑制COX-2。由此可见,COX-2的表达,PGE2的释放和肿瘤细胞的增殖之间有一定的相关性。因此,如选用高选择性COX-2的抑制剂,将对PGE2的释放和肿瘤细胞的增殖产生显著的作用,即对于胃癌的治疗具有潜在的意义。
PMA可通过蛋白激酶C途径而诱导COX-2的表达,使COX2酶活性增加,进一步导致细胞增殖及PGE2释放,其对不表达COX-2的MGC803细胞的效应显著;而对本身可表达COX-2的SGC7901,其作用很微弱甚至表现为抑制。有可能在SGC7901中,COX-2的转录已启动或已处于转录激活状态,此时PMA对其影响的作用很弱。MGC803细胞内与培养上清中PGE2水平的差异悬殊,而SGC7901则不显著,可能为同样的原因,即由于SGC7901中COX-2的转录已经启动,其酶活性催化的PGE2已多数释放至上清中。MGC803不表达COX--2,释放至上清中的PGE2水平则低。这种低水平的PGE2可能为PG产生过程中的另一限速酶COX-1引起。
, http://www.100md.com
有关COX-2对肿瘤免疫的影响,文献报道不多。Urata等〔11〕发现,人胃癌细胞COX-2呈过表达时,可增强淋巴细胞浸润与转移。Tsujii发现〔12〕,结肠癌细胞表达的COX-2可抑制中性粒细胞浸润和巨噬细胞活化,从而导致抗肿瘤免疫监视机能抑制。另外,在非小细胞肺癌中,COX-2阳性的肿瘤细胞可引起细胞因子失衡:即增加巨噬细胞、淋巴细胞产生IL-10,抑制巨噬细胞产生IL-12。IL-12可诱导Th1细胞产生Th1型细胞因子,介导有效的抗肿瘤免疫应答〔8〕。COX-2也可通过PG在肿瘤免疫中发挥其作用:如PGE2可抑制具有免疫调节功能的淋巴因子产生,抑制T细胞和B细胞增殖,T细胞母细胞化及抑制NK细胞、LAK细胞和CTL的细胞毒活性。同时,PGE2还可抑制TNF的产生,诱导IL-10产生。因此,应用环氧化酶抑制剂(如阿斯匹林、苏灵达和消炎痛等)可减低肿瘤引起的免疫抑制作用〔2〕。■
Fig 1 Expression of COX 2 mRNA in tumor cells 1: Negative control;2:fbs; 3:HCC7721; 4:K562; 5:Lo2;6:MGC803; 7:AGS; 8:SGC7901; 9:MKN45;10:100 bp DNA ladder; 11:Caco 2.
, 百拇医药
Fig 2 FCS′ s influence on COX 2 mRNA expression 1: Negative control; 2:SGC7901 10% FCS; 3:AGS 10% FCS; 4:100 bp DNA ladder; 5:AGS DMEM/F12; 6:SGC7901 DMEM/F12.
Fig 3 Immunohistochemical staining of COX 2 (× 40) A: COX 1; B: COX 2; C: Rabbit IgG.
Fig 4 The influence of COX 2 inductor and inhibitor on produ ction of PGE2 from MGC803 cells and in culture supernatant
Fig 5 The influence of indomethacin on release of PGE2 from SGC7901 cells and in culture supernatant
, http://www.100md.com
Fig 6 The influence of indomethacin of different concentrations on SGC7901 cell proliferation and activity 1~ 6: Concentrations of indomathacin are successively 0,2,1,0.5,0.25 and 0.125 μ mol/L.
基金项目:国家自然科学基金资助,No.39870230
作者简介:李玲,女,27岁,硕士生西安市长乐西路15号,Tel.(029)3375229 2000 by J Cell Mol Immunol Press www.immunology. net/jcmi; Email. immuedit@fmmu.edu.cn
参考文献:
, http://www.100md.com 〔1〕 Hinson R, Williams JA, Shacter E. Elevated interleukin 6 is induced by prostaglandin E2 in a murine model of inflammation: possible role of cyclooxygenase 2〔J〕 . Proc Natl Acad Sci USA, 1996; 93(10):4885- 4890.
〔2〕 吴 开 春 , 邓 敬 兰 , 张 学 庸 .胃 癌 组 织 PGE产 生 及 其 对 PHA介 导 的 3H TdR掺 入 的 抑 制 〔J〕 .第 四 军 医 大 学 学 报 , 1988; 9 (5): 295- 296.
〔3〕 Subbaramiah K, Zakim D, Weksler BB, et al. Inhibition of cyclooxygenase: a novel approach to cancer prevention〔J〕 . Proc Soc Exp Biol Med, 1997; 216 (2):201- 210.
, http://www.100md.com
〔4〕 Eberhart CE, Coffey RJ, Radhika A, et al. Up regulation of cyclooxygenase 2 gene expression in human colorectal adenomas and adenocarcinomas〔J〕 . Gastroenterology, 1994; 107(4):1183- 1188.
〔5〕 H, Kawahito Y, Wilder RL, et al. Expression of cyclooxygenase 1 and 2 in human colorectal cancer〔J〕 . Cancer Res, 1995; 55 (17):3785- 3789.
〔6〕 Ristimaki A, Honkanen N, Jankala H, et al. Expression of cyclooxygenase 2 in human gastric carcinoma〔J〕 . Cancer Res, 1997; 57 (7):1276- 1280.
, 百拇医药
〔7〕 Tucker ON, Dannenberg AJ, Yang EK, et al. Cyclooxygenase 2 expression is up regulated in human pancreatic cancer〔J〕 . Cancer Res, 1999; 59 (5): 987- 990.
〔8〕 Huang M, Stolina M, Sharma S, et al. Non small cell lung cancer cyclooxygenase 2 dependent regulation of cytokine balance in lymphocytes and macrophages: up regulation of interleukin 10 and down regulation of interleukin 12 production〔J〕 . Cancer Res, 1998; 58 (7):1208- 1216.
, 百拇医药
〔9〕 Tjandrawinata RR, Dahiya R, Hughes Fulford M. Induction of cyclooxygenase 2 mRNA by prostaglandin E2 in human prostatic carcinoma cells〔J〕 . Br J Cancer, 1997; 75 (8):1111- 1118.
〔10〕 Wong E, Deluca C, Boily C, et al. Characterization of autocrine inducible prostaglandin H synthase 2 (PGHS 2) in human osteos arcoma cells〔J〕 . Inflamm Res, 1997; 46 (1): 51- 56.
〔11〕 Urata H, Kawano S, Tsujii S, et al. Enhanced lymphatic invasion and metastasis with cyclooxygenase 2 overexpression in human gastric carcinoma〔J〕 . Gastroenterology, 1998; 114(4pt2): A798.
〔12〕 Tsujii M, Kawano S, Tsujii S, et al. Effect of cyclooxygenase 2 expression in colon cancer cells on chemotaxis of neutrophils and macrophage〔J〕 . Gastroenterology, 1998; 114(4pt2): A797.
收稿日期:2000-01-02
修回日期:2000-02-16, 百拇医药