��������-3�������ڴ˾��б�����о�
���ߣ������� ����� ��С�� ������ ���ٻ� ������ ����ƽ IJ���� л��
��λ��������(�Ϻ��мƻ�������ѧ�о������Ϻ� 200032)�������(�Ϻ��мƻ�������ѧ�о������Ϻ� 200032)����С��(�㽭��ѧ���ҳ�У������ϵ�� ���� 310029)��������(�Ϻ��мƻ�������ѧ�о������Ϻ� 200032)
�ؼ��ʣ���ZP3��cDNA���˾����������
�й�����ѧ��־000607 ժ Ҫ Ŀ�ģ�ͨ��ԭ�˱���ϵͳ�������ZP����İп�ԭ����������PCR����ɾ����������-3��(pZP3��)cDNA 5'�˷DZ��������˳���������ĸû���5'��СƬ����˫ø��3'�˴�Ƭ����ApaI λ��������ͨ��˫ø��λ�㽫����������Ŀ�Ļ��������pBV221��������PRPL���������εĶ��¡��������� �������������ת�������������յ�����������SDS-PAGE���Ϲ۲쵽61 kD�����쵰�ױ�����������������Western blot ����ʵ�����ܱ���pZP IgGsʶ�𡣽��ۣ����ǻ�����ȻpZP3�����Ŀɻ���Խ����������ƺ����ۻ���pZP3�����ı������硣
, ��Ĵҽҩ
�й�ͼ������ R392. 11
Study on the expression of porcine zona pellucida (ZP)-3�� in a prokaryotic system
XU Wan-Xiang, QIU De-Yi, JIA Xiao-Ming et al
(Shanghai Institute of Planned Parenthood Research, Shanghai 200032)
Abstract Objective:To provide the target antigen for the development of a ZP vaccine by using a prokaryotic expression system. Methods:To obtain non-fusion expression, the fragment (249 bp) from ATG to Apa Isite of pZP3�� cDNA, which was deleted the 5'-terminus non-coding region, was amplified by PCR. The amplified sequenced EcoRI- Apa �� fragment was ligased with the 3'-terminus Apa I -I��Sal I fragment of pZP3�� cDNA from pZ58 plasmid, and then the reconstructed pZP3�� with a complete open reading frame was inserted in frame downstream of the PRPL promoter in a pBV221 vector. Results:After transformation of E.coli BL21 (DE3) and thermo-induction, the pZP3�� protein with 61 kD was expressed at the level of more than 10% of the total cellular proteins and the specific protein band on SDS-PAGE gel could be recognized by anti-porcine ZP IgGs in Western blotting. Conclusion: The availability of native and non-glycosylated pZP3�� protein will help in the development and evaluation of a contraceptive vaccine based on pZP3�� protein.
, ��Ĵҽҩ
Key words Porcine ZP3�� E.coli Gene expression
���鶯��������(ZP)һֱ���������ܾ����������DZ�ڰп�ԭ��Ҳ����ֳ����ѧ���о�����ʶ�𡢽�Ϻʹ����¼��Լ����Ӷ��巴Ӧ����Ҫ����[1��2]��������һ�㲻���봿����Ȼ��ZP���ر�������3�������ǵ�����֣�����谭��ZP�����������ƵĽ��̣������Ծ����ܾ��������ϸ�ڵ�������������Ž�����������ZP��ֵ�3��cDNA��̿�¡����ͼͨ�����̴����������Ҫ��ZP�������Ҳ����ZP�о���һ���ȵ㡣��ǰ�����ѱ�����pZP3�� cDNA ���ںϵ�����ʽ�ڴ˾��б�����о�[3]�����ڽض� ��-��������ø���ʱ��о��Ķ����÷��ǻ���pZP3���ںϵ��ķ�����(Լ102 kD)�ϴ�����δ�ﵽ���ڴ���ˮƽ��pZP3������ֱ�ӱ���;����
1 �����뷽��
, http://www.100md.com 1.1 �����;��� pBluescript KS (pBS) ����Ϊ�������壬E. coli BL21(DE3)Ϊ����ת���������PRPL�����ӱ�������pBV220ΪԤ��ҽѧ��ѧԺ��������������ڹ�����PBV220����������pBV221���п�Ժ�������﷽�鲩ʿ��������ȫ��pZP3�� cDNA ��pZ58����(pBS/pZP3��)������Zonagen Inc. Harris��ʿ������
1.2 �Լ���ø EcoRI��ApaI��SalI����������ø�Լ�T4 DNA ����ø����Boeringer Mannheim��˾��Taq DNA�ۺ�ø�����Ϻ����չ�˾������͵����ʷ��������Լ�����IgG/HRP���Ի������﹤�̹�˾�����������Լ���Ϊ������������
1.3 �ÿ���ZP IgGs�Ʊ� ������[3,4]��
1.4 PCR���� ���ݷ�����pZP3�� cDNA���м�PCR�������ԭ�����һ��PCR����[5]����������P1��5'-GGGAATTCATGTGGTTGCGGCCGTCC-3' (ATGǰ����EcoRIλ��)����������P2��5'-CTCCCATGGAGCTGCCTGGG-3' (3'�˺����ApaIλ��)����pZ58����Ϊģ�����PCR��������Ӧ�������£�94�棬1 min;55 ��, 40 s; 72 ��, 30 s, ��25��ѭ����
, ��Ĵҽҩ
1.5 PCR�����DNA������� PCR ����(271 bp)��EcoRI��ApaI˫ø�е�Ƭ��(pZP3��', 249 bp)���������pBS�������塣�������װ�ɸѡ�������¡��������������DNA��������ȡpBS/pZP3��'����, ��ABI373A��DNA�������Ͻ���������ӫ����DNA�����Զ�������
1.6 ɾ��5'�˷DZ�������pZP3��* cDNA �Լ�pBV220- pZP3��*��pBV221- pZP3��*������������Ĺ��� ɾ��5'�˷DZ�������pZP3��' (EcoRI-ApaI)СƬ����˫ø����pZ58������pZP3��" (ApaI-SalI)��Ƭ��(1 445 bp, ��ԭcDNA 3'��EcoRIλ����pBS���¡��SalIλ��֮���27 bp)��ApaIλ��������Ȼ�˶����������µ�pZP3��* cDNA(1 694 bp) �����������pBV����������ø�С�Ƭ�λ��ա����ӡ�ת����������ȡ�͵�Ӿ�����Ȳ�������������[6]����Ӧ�Լ���˵������С�
, ��Ĵҽҩ
1.7 ���̾�BL21/pBV221- pZP3��*�����յ����� ��ת��������BL21(DE3)/pBV221- pZP3��*���̾������䵥��������ڼ��백����ù��(Ap)��3 ml LB����Һ����30�{C��������ҹ�����ճ���1/50 (V/V)�ı���ת�����Ap��50 ml LB ����Һ����30�{C��������2��3 h, ����Һ��A600����ȴﵽ0.7����ʱ������42 �� ��������4 h���ռ����塣
1.8 ��ZP3�����ױ�������SDS-PAGE�����͵���ӡ������ ������[3-6]��
2 ���
2.1 pZP3�� cDNA 5'�˷DZ�������ɾ�� ͨ��ԭ������ֱ�ӱ���Ŀ�ĵ��ף����ڱ��������ϵ�SD������Ŀ�Ļ���ATG֮���������ơ�ѡ��PCR����ɾ��pZP3�� cDNA 5'�˵ķDZ�����(33 bp)������Ŀ�Ļ���1 700 bp[3,5], ����Ϊ��ֹPCR�����п��ܲ����ļ��ͻ�䣬�������˸û��������ʼ����ATG��ǰ��ApaI��һø��λ��Ķ�Ƭ��(271 bp)����˫ø�к��EcoRI-ApaIƬ��(pZP3��', 249 bp)���������pBS��������DNA�������(ͼ1)��ø�м��������ͼ2��Ȼ��ø�д�������pBS-pZP3��'������EcoRI-ApaI pZP3��' ��Ƭ���Լ�����pZ58������pZP3�� cDNA 3'�˵�ApaI-SalI��Ƭ��(pZP3��", 1 445 bp)��ͨ��DNA����ø��ApaIλ��ʹ֮�����ӣ����չ�����ɾ����ԭ��¡����5'�˴ַDZ�������pZP3��* cDNA��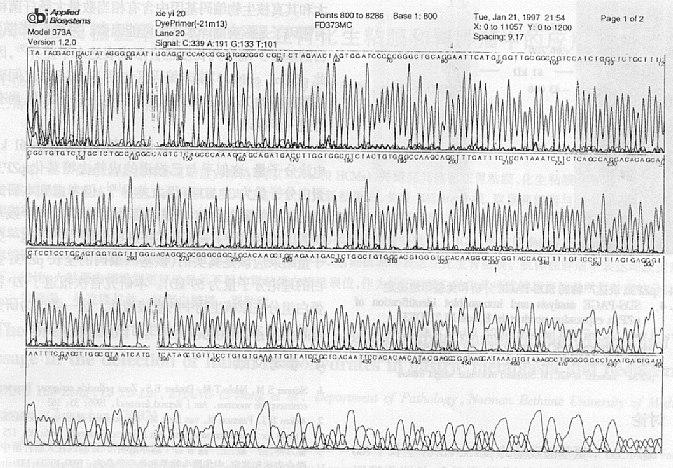
, http://www.100md.com
ͼ1 pZP3������5'��PCRƬ�ε�DNA����ͼ��
Fig.1 Sequencing result for the PCR product of pZP3�� cDNA
Note: "��" and "��", indicate the sequenced PCR fragment from EcoRI to ApaI site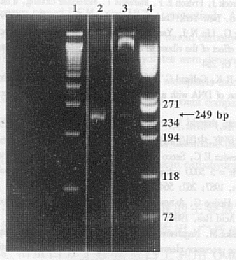
ͼ2 PCRƬ�ε�5%�۱�ϩ����������Ӿ
Fig.2 5% PAGE analysis of the PCR fragment
Note�� 1. 100 bp ladder��2. The PCR fragment digested by EcoRI and ApaI��
, http://www.100md.com
3. The PCR fragment from pBV221- pZP3�� after digestion by EcoRI-ApaI��
4. The �� x 174/Hae III marker
2.2 pBV220- pZP3��* ��pBV221-pZP3��*������������Ĺ��� ����֬��������Ӿ���յľ���EcoRI��SalIճ��ĩ�˵�pZP3��'-pZP3������Ƭ�� pZP3��*����DNA����ø�������£��������������pBV220��pBV221��������PRPL���������ζ��¡����EcoRI��SalIλ�㡣���ӷ�ӦҺת���˾�BL21(DE3)���������ڼ��백����ù��(Ap)��LB����ƽ������ѡ���ɵ������������������������DNA�����Լ��г���������������ø�м���(ͼ3)���Ӷ�ȷ��BL21(DE3)/pBV220- pZP3��*��BL21(DE3)/pBV221- pZP3��*���̾���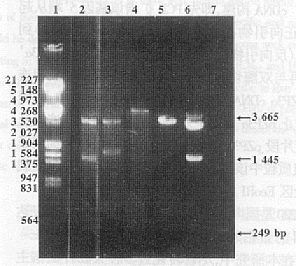
, http://www.100md.com
ͼ3 ��������pBV221- pZP3����ø�м���
Fig.3 1.2% agarose gel analysis of restricted fragments from pBV221-pZP3��* recombinant plasmid
Note: 1.�� DNA /EcoRI+HidIII marker��2. pBV221-pZP3��* digested by EcoRI+ApaI��
3. pBV221-pZP3��*digested by EcoRI��4. pBV221-pZP3�� digested by SalI��
5. pBV221 digested by EcoRI��6. PBS/ pZP3�� (pZ58) digested by ApaI+SalI��
, ��Ĵҽҩ
7. The PCR fragment digested by EcoRI+ApaI
2.3 pZP3�������ڴ˾��е�ֱ�ӱ��� pBV221��pBV220��������������[7], �������pBV221 PRPL���������ε�Ŀ�Ļ���ֱ�ӱ������¶ȱ仯���ء��ռ������յ�BL21(DE3)/pBV221- pZP3��*���̾����徭�ѽ�Һ������ȡ����12% SDS-PAGE�����������61 kD���ɷ����б��ڶ��վ����״���pZP3��������������(ͼ4)���������������ɨ������ʾ�˱��ﵰ��ռ�����ܵ���10%���ϡ�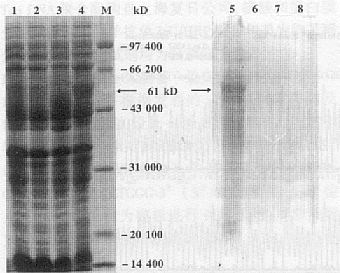
ͼ4 pZP3����������SDS-PAGE����������ӡ������
Fig.4 SDS-PAGE analysis and immunoblot identification of pZP3��
, http://www.100md.com
expression product in E.coli BL21(DE3)
Note: 1 and 8. E. coli BL21 harboring pBV221 induced at 42�{C��
2 and 7. E. coli BL21 induced at 42�{C��3 and 6. E. coli BL21 harboring
pBV221- pZP3��* cultured at 30�棻4 and 5. E. coli BL21 harboring
pBV221- pZP3��* induced at 42�{C��M.Standard molecular weight marker
2.4 pZP3�����ױ�����������ԭ�Լ��� Ϊ��֤ʵSDS-PAGE�������б��ڶ��յ�������������pZP3�����ױ��������ǽ�����Western blot�������ڽ�������Ʒ��ͬ����һ����������Ӿ����תӡ��������ά��Ĥ�Ϻ��ú�pZP3��������ÿ���ZP IgGs ������IgGs/HRP�ֱ����һ���Ͷ�����Ӧ������ڹ��̾��յ�����ӡ�����������ؿ�����SDS-PAGE��������������ƽ��һ�µ����Է�Ӧ��ɫ������3�ֶ��վ����״���ӡ�����������Է�Ӧ�ź�(ͼ4)�������pZP3��������BL21(DE3)/pBV221-pZP3��*���̾���ȷʵ����˱��
, ��Ĵҽҩ
3 ����
����536��������л���pZP3�� cDNAȫ��1 700 bp�������û���5'��38 bp�DZ�������3'�˷DZ�����[3,5]�����ǵ�PCR����������һ���Ĵ�����[8]���Ҳ���һ�����Լ1 600 bp����Ƭ�ε�DNA������������������ڱ��о��з��������ɾ��5'�˷DZ���33 bp��pZP3��* cDNA����������PCR�����û���5'�˴���ʼ���� (������������֮ǰ��������EcoRIλ��)��ApaIλ��(������������ڽ�������������)��pZP3��'��Ƭ�Σ�����˫ø��3'��ApaI-SalI pZP3��"��Ƭ�������ӡ�ԭpZP3�� cDNA 3'�˿�¡λ��Ҳ��EcoRIλ�㣬���Ϊ�˷�ֹpZP3��'��pZP3��"��EcoRIλ��������Լ���������Ƭ��pZP3��*�������pBV���壬����ѡ����pZ58���������иû���3'��EcoRIλ���pBS��¡������¡��EcoRI���SalIλ��(���27 bps)��
, ��Ĵҽҩ
pBV220�ǹ���Ӧ�ù㷺�����յ�������������������ʱĿ�ĵ��ױ������ɴ�����ܵ���30%��70%�����ڱ��о��У�����ת�������ִ˾���������pBV220- pZP3��*�������������δ�γ���SDS-PAGE�пɱ������쵰�ױ��pZP3��*������������������pBV221�������BL21(DE3)�������л���˸���10%����������Ȼ��һ�������������룬��������ǰԼ5%���ںϵ���(102 kD)����ˮƽ���[3]��ȥ���ںϵ�����Լռһ��л��Ľض� ��-��������ø��pZP3��Ŀ�ĵ��ױ��ﻹ������˽�4��������Ŀ�ĵ��ױ�����Խϵͣ��Ʋ�pZP3�����ױ����������ϴ������������������к����൱�����˾�ϡ����������Ӱ�췭��ˮƽ�Ŀ������ء������б���˵Ŀ�Ļ���3'�����г���Ҳ���γɱ���ˮƽ����[9]�����pZP3��* cDNA 3'�˷DZ���������EcoRI-SalI֮���������ڵ�76 bp�DZ������Ƿ�Ҳ��һ��ԭ�����д���һ��̽�֡�
, http://www.100md.com
��SDS-PAGE��pZP3�����ױ���������ʾ61 kD��Ӿ�����������ƺ����ѱ����ij���ȥ�ǻ���pZP3����������Ϊ37 kD�кܴ����[10]�������ǵ����о������Ŀ�ĵ���21���л��ź��ĺ�C��74���л���Ĥ��(���쵰��һ�㲻������Щ�Ķ�[11-13]), ��һ���������Ӧ������ʵ��(�û�������ȫ��pZP3���������۷�����Ϊ59 kD)�����о��״α�����ZP�ǵ��������ԭ�������е�ֱ�ӱ����һ����Ϊ�о�ȥ�ǻ���ZP������ֵĿ���������춨�˻�����
���Ҽƻ�����ҩ���ص�ʵ��������
����飺�����飬50�꣬���о�Ա����Ҫ������ֳ����ѧ�о���
л�㣬35�꣬���ڣ���Ҫ���·�������ѧ�о�
���ٻ�(������ѧ������ѧѧԺ�Ŵ��о����Ŵ����̹����ص�ʵ���ң� �Ϻ� 200433)
, ��Ĵҽҩ
������(��������ҽѧԺ�� �ߺ� 241001)
����ƽ(������ѧ������ѧѧԺ�Ŵ��о����Ŵ����̹����ص�ʵ���ң� �Ϻ� 200433)
IJ����(������ѧ������ѧѧԺ�Ŵ��о����Ŵ����̹����ص�ʵ���ң� �Ϻ� 200433)
л��(������ѧ������ѧѧԺ�Ŵ��о����Ŵ����̹����ص�ʵ���ң� �Ϻ� 200433)
�����
1��Skinner S M, Ndolo T M, Dunber B S. Zona pellucida antigens: Targets for contraceptive vaccines. Am J Reprod Immunol, 1996; 35: 163
, ��Ĵҽҩ 2��Aitken R J, Paterson M, van Duin M. The potential of the zona pellucida as a target for immunocontraception. Am J Reprod Immunol, 1996; 35: 175
3�������飬л �㣬�� �� et al. ��������-3�������ڴ˾��е��ںϱ���ͼ���.�л�����ѧ������ѧ��־�� 1999��19(2)��137
4�������飬����壬�� �� et al. ��������-3���ںϵ�����E. coli�еı���ͼ���.�й�����ѧ��־�� 1998��14(6): 420
5��Yurewicz E C, Hibler D, Fontenot G K et al. Nucleotide sequence of cDNA encoding ZP3��, a sperm-binding glycoprotein from zona pellucida of pig oocyte. Biochim Biophys Acta, 1993; 1174: 211
, ��Ĵҽҩ
6��Sambrook J, Fritsch E F, Maniatis T. Molecular cloning, a laboratory manual 2nd ed. New York��Cold Spring Harbor Laboratory Press��1989��80
7��Wang G, Lin N J, Yang K Y. High-level expression of prochymosin in E. coli: effect of the ribosome binding site. Protein Expression and Purification, 1995; 6: 284
8��Saiki R K, Gelfand G T, Stoffel S J et al. Primer-directed enzymatic amplification of DNA with a thermostable DNA polymerase. Science, 1988; 239: 487
, ��Ĵҽҩ
9���� ����¬���ɣ��� �� et al. ������ѪС���������ڴ˾��б�����о�.�й����ﻯѧ���������ѧ���� 1999��15(1): 19
10��Yurewicz E C, Sacco A G, Subramnian M G. Structural characterization of the Mr=5 5000 antigen (ZP3) of porcine oocyte zona pellucida. J Biol Chem, 1987; 262: 564
11��Von Heijne G. A new method for predicting signal sequence cleavage site. Nuc Acid Res, 1986; 14: 4683
12��Hosaka M, Nagahama M, Kim W et al. Arg-X-Lys/Arg-Arg motif as a signal for precursor cleavage-catalyzed by furin within the constitutive secretary pathway. J Biol Chem, 1991; 266: 12127
13��Kaul R, Afzalpurkar A, Gupta S K. Expression of bonnet monkey (Macaca radiata) zona pellucida-3 (ZP3) in a prokaryotic system and its immunogenicity. Mol Reprod Dev, 1997; 47: 140
�ո�1999-09-16 ��2000-02-12, http://www.100md.com
��λ��������(�Ϻ��мƻ�������ѧ�о������Ϻ� 200032)�������(�Ϻ��мƻ�������ѧ�о������Ϻ� 200032)����С��(�㽭��ѧ���ҳ�У������ϵ�� ���� 310029)��������(�Ϻ��мƻ�������ѧ�о������Ϻ� 200032)
�ؼ��ʣ���ZP3��cDNA���˾����������
�й�����ѧ��־000607 ժ Ҫ Ŀ�ģ�ͨ��ԭ�˱���ϵͳ�������ZP����İп�ԭ����������PCR����ɾ����������-3��(pZP3��)cDNA 5'�˷DZ��������˳���������ĸû���5'��СƬ����˫ø��3'�˴�Ƭ����ApaI λ��������ͨ��˫ø��λ�㽫����������Ŀ�Ļ��������pBV221��������PRPL���������εĶ��¡��������� �������������ת�������������յ�����������SDS-PAGE���Ϲ۲쵽61 kD�����쵰�ױ�����������������Western blot ����ʵ�����ܱ���pZP IgGsʶ�𡣽��ۣ����ǻ�����ȻpZP3�����Ŀɻ���Խ����������ƺ����ۻ���pZP3�����ı������硣
, ��Ĵҽҩ
�й�ͼ������ R392. 11
Study on the expression of porcine zona pellucida (ZP)-3�� in a prokaryotic system
XU Wan-Xiang, QIU De-Yi, JIA Xiao-Ming et al
(Shanghai Institute of Planned Parenthood Research, Shanghai 200032)
Abstract Objective:To provide the target antigen for the development of a ZP vaccine by using a prokaryotic expression system. Methods:To obtain non-fusion expression, the fragment (249 bp) from ATG to Apa Isite of pZP3�� cDNA, which was deleted the 5'-terminus non-coding region, was amplified by PCR. The amplified sequenced EcoRI- Apa �� fragment was ligased with the 3'-terminus Apa I -I��Sal I fragment of pZP3�� cDNA from pZ58 plasmid, and then the reconstructed pZP3�� with a complete open reading frame was inserted in frame downstream of the PRPL promoter in a pBV221 vector. Results:After transformation of E.coli BL21 (DE3) and thermo-induction, the pZP3�� protein with 61 kD was expressed at the level of more than 10% of the total cellular proteins and the specific protein band on SDS-PAGE gel could be recognized by anti-porcine ZP IgGs in Western blotting. Conclusion: The availability of native and non-glycosylated pZP3�� protein will help in the development and evaluation of a contraceptive vaccine based on pZP3�� protein.
, ��Ĵҽҩ
Key words Porcine ZP3�� E.coli Gene expression
���鶯��������(ZP)һֱ���������ܾ����������DZ�ڰп�ԭ��Ҳ����ֳ����ѧ���о�����ʶ�𡢽�Ϻʹ����¼��Լ����Ӷ��巴Ӧ����Ҫ����[1��2]��������һ�㲻���봿����Ȼ��ZP���ر�������3�������ǵ�����֣�����谭��ZP�����������ƵĽ��̣������Ծ����ܾ��������ϸ�ڵ�������������Ž�����������ZP��ֵ�3��cDNA��̿�¡����ͼͨ�����̴����������Ҫ��ZP�������Ҳ����ZP�о���һ���ȵ㡣��ǰ�����ѱ�����pZP3�� cDNA ���ںϵ�����ʽ�ڴ˾��б�����о�[3]�����ڽض� ��-��������ø���ʱ��о��Ķ����÷��ǻ���pZP3���ںϵ��ķ�����(Լ102 kD)�ϴ�����δ�ﵽ���ڴ���ˮƽ��pZP3������ֱ�ӱ���;����
1 �����뷽��
, http://www.100md.com 1.1 �����;��� pBluescript KS (pBS) ����Ϊ�������壬E. coli BL21(DE3)Ϊ����ת���������PRPL�����ӱ�������pBV220ΪԤ��ҽѧ��ѧԺ��������������ڹ�����PBV220����������pBV221���п�Ժ�������﷽�鲩ʿ��������ȫ��pZP3�� cDNA ��pZ58����(pBS/pZP3��)������Zonagen Inc. Harris��ʿ������
1.2 �Լ���ø EcoRI��ApaI��SalI����������ø�Լ�T4 DNA ����ø����Boeringer Mannheim��˾��Taq DNA�ۺ�ø�����Ϻ����չ�˾������͵����ʷ��������Լ�����IgG/HRP���Ի������﹤�̹�˾�����������Լ���Ϊ������������
1.3 �ÿ���ZP IgGs�Ʊ� ������[3,4]��
1.4 PCR���� ���ݷ�����pZP3�� cDNA���м�PCR�������ԭ�����һ��PCR����[5]����������P1��5'-GGGAATTCATGTGGTTGCGGCCGTCC-3' (ATGǰ����EcoRIλ��)����������P2��5'-CTCCCATGGAGCTGCCTGGG-3' (3'�˺����ApaIλ��)����pZ58����Ϊģ�����PCR��������Ӧ�������£�94�棬1 min;55 ��, 40 s; 72 ��, 30 s, ��25��ѭ����
, ��Ĵҽҩ
1.5 PCR�����DNA������� PCR ����(271 bp)��EcoRI��ApaI˫ø�е�Ƭ��(pZP3��', 249 bp)���������pBS�������塣�������װ�ɸѡ�������¡��������������DNA��������ȡpBS/pZP3��'����, ��ABI373A��DNA�������Ͻ���������ӫ����DNA�����Զ�������
1.6 ɾ��5'�˷DZ�������pZP3��* cDNA �Լ�pBV220- pZP3��*��pBV221- pZP3��*������������Ĺ��� ɾ��5'�˷DZ�������pZP3��' (EcoRI-ApaI)СƬ����˫ø����pZ58������pZP3��" (ApaI-SalI)��Ƭ��(1 445 bp, ��ԭcDNA 3'��EcoRIλ����pBS���¡��SalIλ��֮���27 bp)��ApaIλ��������Ȼ�˶����������µ�pZP3��* cDNA(1 694 bp) �����������pBV����������ø�С�Ƭ�λ��ա����ӡ�ת����������ȡ�͵�Ӿ�����Ȳ�������������[6]����Ӧ�Լ���˵������С�
, ��Ĵҽҩ
1.7 ���̾�BL21/pBV221- pZP3��*�����յ����� ��ת��������BL21(DE3)/pBV221- pZP3��*���̾������䵥��������ڼ��백����ù��(Ap)��3 ml LB����Һ����30�{C��������ҹ�����ճ���1/50 (V/V)�ı���ת�����Ap��50 ml LB ����Һ����30�{C��������2��3 h, ����Һ��A600����ȴﵽ0.7����ʱ������42 �� ��������4 h���ռ����塣
1.8 ��ZP3�����ױ�������SDS-PAGE�����͵���ӡ������ ������[3-6]��
2 ���
2.1 pZP3�� cDNA 5'�˷DZ�������ɾ�� ͨ��ԭ������ֱ�ӱ���Ŀ�ĵ��ף����ڱ��������ϵ�SD������Ŀ�Ļ���ATG֮���������ơ�ѡ��PCR����ɾ��pZP3�� cDNA 5'�˵ķDZ�����(33 bp)������Ŀ�Ļ���1 700 bp[3,5], ����Ϊ��ֹPCR�����п��ܲ����ļ��ͻ�䣬�������˸û��������ʼ����ATG��ǰ��ApaI��һø��λ��Ķ�Ƭ��(271 bp)����˫ø�к��EcoRI-ApaIƬ��(pZP3��', 249 bp)���������pBS��������DNA�������(ͼ1)��ø�м��������ͼ2��Ȼ��ø�д�������pBS-pZP3��'������EcoRI-ApaI pZP3��' ��Ƭ���Լ�����pZ58������pZP3�� cDNA 3'�˵�ApaI-SalI��Ƭ��(pZP3��", 1 445 bp)��ͨ��DNA����ø��ApaIλ��ʹ֮�����ӣ����չ�����ɾ����ԭ��¡����5'�˴ַDZ�������pZP3��* cDNA��
, http://www.100md.com
ͼ1 pZP3������5'��PCRƬ�ε�DNA����ͼ��
Fig.1 Sequencing result for the PCR product of pZP3�� cDNA
Note: "��" and "��", indicate the sequenced PCR fragment from EcoRI to ApaI site
ͼ2 PCRƬ�ε�5%�۱�ϩ����������Ӿ
Fig.2 5% PAGE analysis of the PCR fragment
Note�� 1. 100 bp ladder��2. The PCR fragment digested by EcoRI and ApaI��
, http://www.100md.com
3. The PCR fragment from pBV221- pZP3�� after digestion by EcoRI-ApaI��
4. The �� x 174/Hae III marker
2.2 pBV220- pZP3��* ��pBV221-pZP3��*������������Ĺ��� ����֬��������Ӿ���յľ���EcoRI��SalIճ��ĩ�˵�pZP3��'-pZP3������Ƭ�� pZP3��*����DNA����ø�������£��������������pBV220��pBV221��������PRPL���������ζ��¡����EcoRI��SalIλ�㡣���ӷ�ӦҺת���˾�BL21(DE3)���������ڼ��백����ù��(Ap)��LB����ƽ������ѡ���ɵ������������������������DNA�����Լ��г���������������ø�м���(ͼ3)���Ӷ�ȷ��BL21(DE3)/pBV220- pZP3��*��BL21(DE3)/pBV221- pZP3��*���̾���
, http://www.100md.com
ͼ3 ��������pBV221- pZP3����ø�м���
Fig.3 1.2% agarose gel analysis of restricted fragments from pBV221-pZP3��* recombinant plasmid
Note: 1.�� DNA /EcoRI+HidIII marker��2. pBV221-pZP3��* digested by EcoRI+ApaI��
3. pBV221-pZP3��*digested by EcoRI��4. pBV221-pZP3�� digested by SalI��
5. pBV221 digested by EcoRI��6. PBS/ pZP3�� (pZ58) digested by ApaI+SalI��
, ��Ĵҽҩ
7. The PCR fragment digested by EcoRI+ApaI
2.3 pZP3�������ڴ˾��е�ֱ�ӱ��� pBV221��pBV220��������������[7], �������pBV221 PRPL���������ε�Ŀ�Ļ���ֱ�ӱ������¶ȱ仯���ء��ռ������յ�BL21(DE3)/pBV221- pZP3��*���̾����徭�ѽ�Һ������ȡ����12% SDS-PAGE�����������61 kD���ɷ����б��ڶ��վ����״���pZP3��������������(ͼ4)���������������ɨ������ʾ�˱��ﵰ��ռ�����ܵ���10%���ϡ�
ͼ4 pZP3����������SDS-PAGE����������ӡ������
Fig.4 SDS-PAGE analysis and immunoblot identification of pZP3��
, http://www.100md.com
expression product in E.coli BL21(DE3)
Note: 1 and 8. E. coli BL21 harboring pBV221 induced at 42�{C��
2 and 7. E. coli BL21 induced at 42�{C��3 and 6. E. coli BL21 harboring
pBV221- pZP3��* cultured at 30�棻4 and 5. E. coli BL21 harboring
pBV221- pZP3��* induced at 42�{C��M.Standard molecular weight marker
2.4 pZP3�����ױ�����������ԭ�Լ��� Ϊ��֤ʵSDS-PAGE�������б��ڶ��յ�������������pZP3�����ױ��������ǽ�����Western blot�������ڽ�������Ʒ��ͬ����һ����������Ӿ����תӡ��������ά��Ĥ�Ϻ��ú�pZP3��������ÿ���ZP IgGs ������IgGs/HRP�ֱ����һ���Ͷ�����Ӧ������ڹ��̾��յ�����ӡ�����������ؿ�����SDS-PAGE��������������ƽ��һ�µ����Է�Ӧ��ɫ������3�ֶ��վ����״���ӡ�����������Է�Ӧ�ź�(ͼ4)�������pZP3��������BL21(DE3)/pBV221-pZP3��*���̾���ȷʵ����˱��
, ��Ĵҽҩ
3 ����
����536��������л���pZP3�� cDNAȫ��1 700 bp�������û���5'��38 bp�DZ�������3'�˷DZ�����[3,5]�����ǵ�PCR����������һ���Ĵ�����[8]���Ҳ���һ�����Լ1 600 bp����Ƭ�ε�DNA������������������ڱ��о��з��������ɾ��5'�˷DZ���33 bp��pZP3��* cDNA����������PCR�����û���5'�˴���ʼ���� (������������֮ǰ��������EcoRIλ��)��ApaIλ��(������������ڽ�������������)��pZP3��'��Ƭ�Σ�����˫ø��3'��ApaI-SalI pZP3��"��Ƭ�������ӡ�ԭpZP3�� cDNA 3'�˿�¡λ��Ҳ��EcoRIλ�㣬���Ϊ�˷�ֹpZP3��'��pZP3��"��EcoRIλ��������Լ���������Ƭ��pZP3��*�������pBV���壬����ѡ����pZ58���������иû���3'��EcoRIλ���pBS��¡������¡��EcoRI���SalIλ��(���27 bps)��
, ��Ĵҽҩ
pBV220�ǹ���Ӧ�ù㷺�����յ�������������������ʱĿ�ĵ��ױ������ɴ�����ܵ���30%��70%�����ڱ��о��У�����ת�������ִ˾���������pBV220- pZP3��*�������������δ�γ���SDS-PAGE�пɱ������쵰�ױ��pZP3��*������������������pBV221�������BL21(DE3)�������л���˸���10%����������Ȼ��һ�������������룬��������ǰԼ5%���ںϵ���(102 kD)����ˮƽ���[3]��ȥ���ںϵ�����Լռһ��л��Ľض� ��-��������ø��pZP3��Ŀ�ĵ��ױ��ﻹ������˽�4��������Ŀ�ĵ��ױ�����Խϵͣ��Ʋ�pZP3�����ױ����������ϴ������������������к����൱�����˾�ϡ����������Ӱ�췭��ˮƽ�Ŀ������ء������б���˵Ŀ�Ļ���3'�����г���Ҳ���γɱ���ˮƽ����[9]�����pZP3��* cDNA 3'�˷DZ���������EcoRI-SalI֮���������ڵ�76 bp�DZ������Ƿ�Ҳ��һ��ԭ�����д���һ��̽�֡�
, http://www.100md.com
��SDS-PAGE��pZP3�����ױ���������ʾ61 kD��Ӿ�����������ƺ����ѱ����ij���ȥ�ǻ���pZP3����������Ϊ37 kD�кܴ����[10]�������ǵ����о������Ŀ�ĵ���21���л��ź��ĺ�C��74���л���Ĥ��(���쵰��һ�㲻������Щ�Ķ�[11-13]), ��һ���������Ӧ������ʵ��(�û�������ȫ��pZP3���������۷�����Ϊ59 kD)�����о��״α�����ZP�ǵ��������ԭ�������е�ֱ�ӱ����һ����Ϊ�о�ȥ�ǻ���ZP������ֵĿ���������춨�˻�����
���Ҽƻ�����ҩ���ص�ʵ��������
����飺�����飬50�꣬���о�Ա����Ҫ������ֳ����ѧ�о���
л�㣬35�꣬���ڣ���Ҫ���·�������ѧ�о�
���ٻ�(������ѧ������ѧѧԺ�Ŵ��о����Ŵ����̹����ص�ʵ���ң� �Ϻ� 200433)
, ��Ĵҽҩ
������(��������ҽѧԺ�� �ߺ� 241001)
����ƽ(������ѧ������ѧѧԺ�Ŵ��о����Ŵ����̹����ص�ʵ���ң� �Ϻ� 200433)
IJ����(������ѧ������ѧѧԺ�Ŵ��о����Ŵ����̹����ص�ʵ���ң� �Ϻ� 200433)
л��(������ѧ������ѧѧԺ�Ŵ��о����Ŵ����̹����ص�ʵ���ң� �Ϻ� 200433)
�����
1��Skinner S M, Ndolo T M, Dunber B S. Zona pellucida antigens: Targets for contraceptive vaccines. Am J Reprod Immunol, 1996; 35: 163
, ��Ĵҽҩ 2��Aitken R J, Paterson M, van Duin M. The potential of the zona pellucida as a target for immunocontraception. Am J Reprod Immunol, 1996; 35: 175
3�������飬л �㣬�� �� et al. ��������-3�������ڴ˾��е��ںϱ���ͼ���.�л�����ѧ������ѧ��־�� 1999��19(2)��137
4�������飬����壬�� �� et al. ��������-3���ںϵ�����E. coli�еı���ͼ���.�й�����ѧ��־�� 1998��14(6): 420
5��Yurewicz E C, Hibler D, Fontenot G K et al. Nucleotide sequence of cDNA encoding ZP3��, a sperm-binding glycoprotein from zona pellucida of pig oocyte. Biochim Biophys Acta, 1993; 1174: 211
, ��Ĵҽҩ
6��Sambrook J, Fritsch E F, Maniatis T. Molecular cloning, a laboratory manual 2nd ed. New York��Cold Spring Harbor Laboratory Press��1989��80
7��Wang G, Lin N J, Yang K Y. High-level expression of prochymosin in E. coli: effect of the ribosome binding site. Protein Expression and Purification, 1995; 6: 284
8��Saiki R K, Gelfand G T, Stoffel S J et al. Primer-directed enzymatic amplification of DNA with a thermostable DNA polymerase. Science, 1988; 239: 487
, ��Ĵҽҩ
9���� ����¬���ɣ��� �� et al. ������ѪС���������ڴ˾��б�����о�.�й����ﻯѧ���������ѧ���� 1999��15(1): 19
10��Yurewicz E C, Sacco A G, Subramnian M G. Structural characterization of the Mr=5 5000 antigen (ZP3) of porcine oocyte zona pellucida. J Biol Chem, 1987; 262: 564
11��Von Heijne G. A new method for predicting signal sequence cleavage site. Nuc Acid Res, 1986; 14: 4683
12��Hosaka M, Nagahama M, Kim W et al. Arg-X-Lys/Arg-Arg motif as a signal for precursor cleavage-catalyzed by furin within the constitutive secretary pathway. J Biol Chem, 1991; 266: 12127
13��Kaul R, Afzalpurkar A, Gupta S K. Expression of bonnet monkey (Macaca radiata) zona pellucida-3 (ZP3) in a prokaryotic system and its immunogenicity. Mol Reprod Dev, 1997; 47: 140
�ո�1999-09-16 ��2000-02-12, http://www.100md.com